
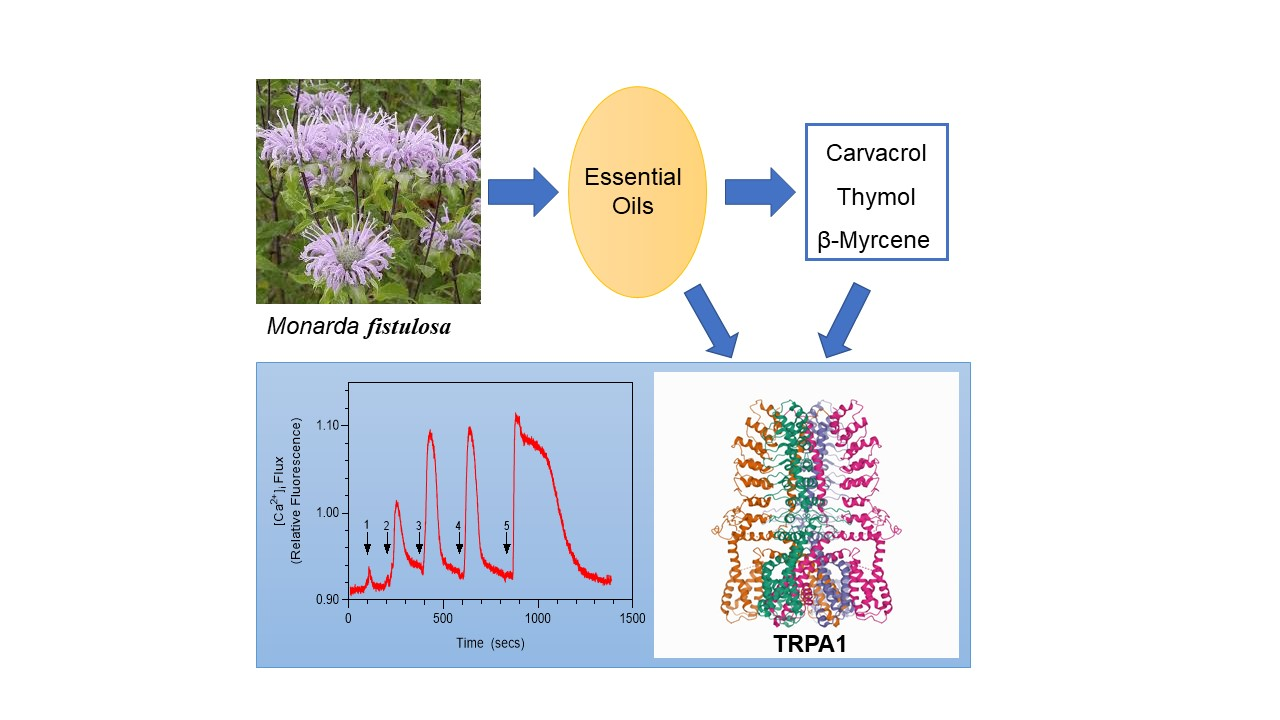
Essential Oils from Monarda fistulosa: Chemical Composition and Activation of Transient Receptor Potential A1 (TRPA1) Channels

 ,
, 
 ,
, 
Abstract
:
1. Introduction
2. Results
2.1. Essential Oil Composition
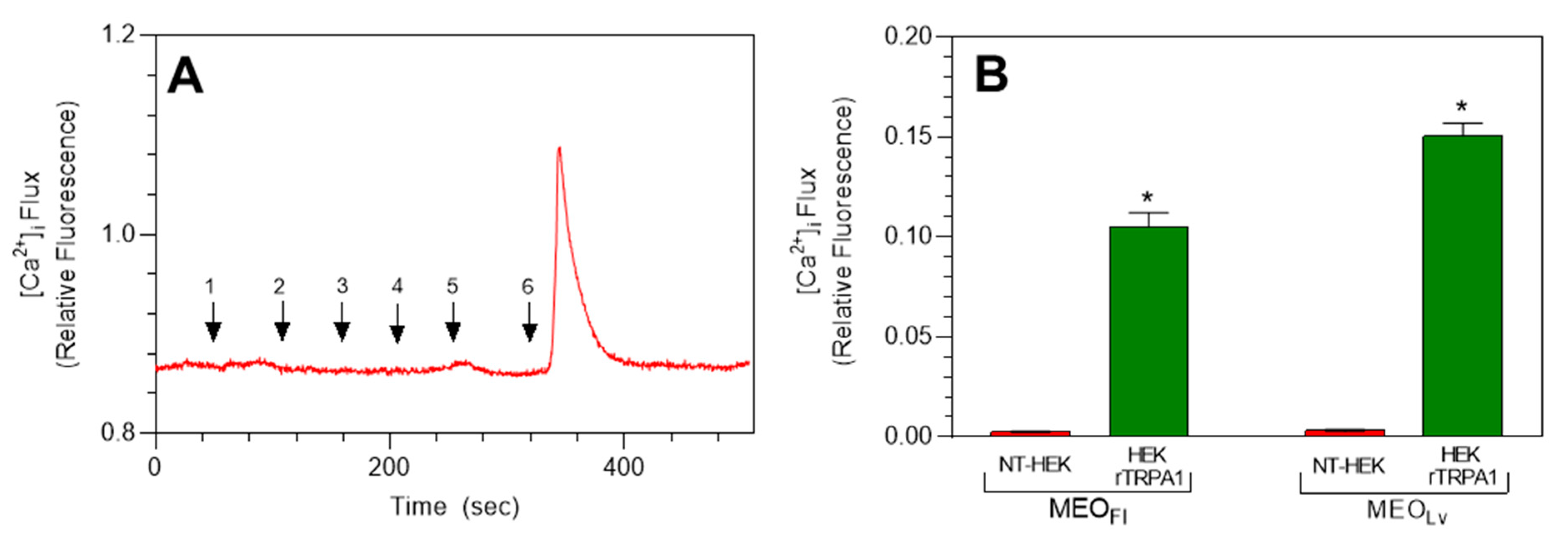
2.2. Effect of M. fistulosa Essential Oils on TRP Ion Channels
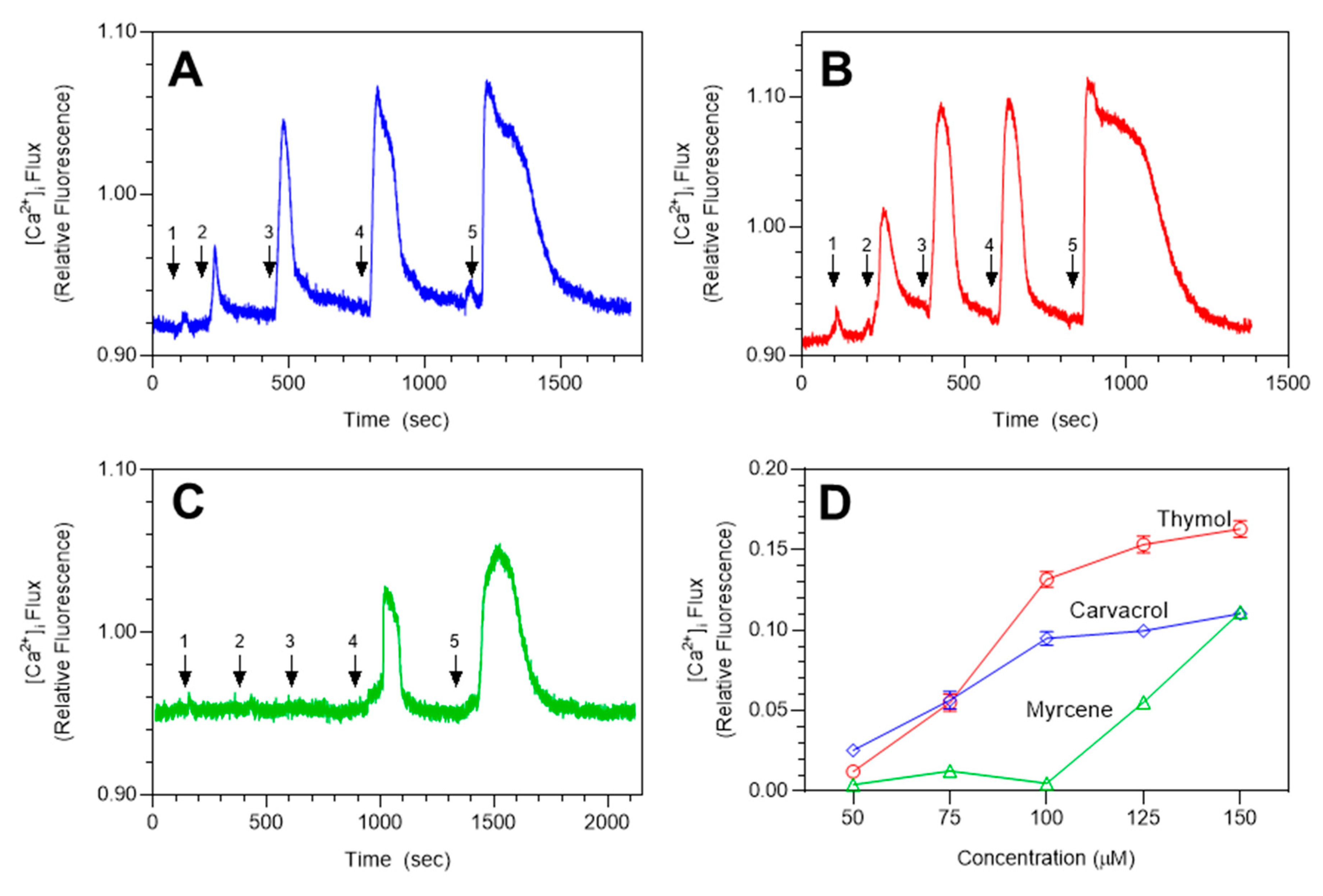
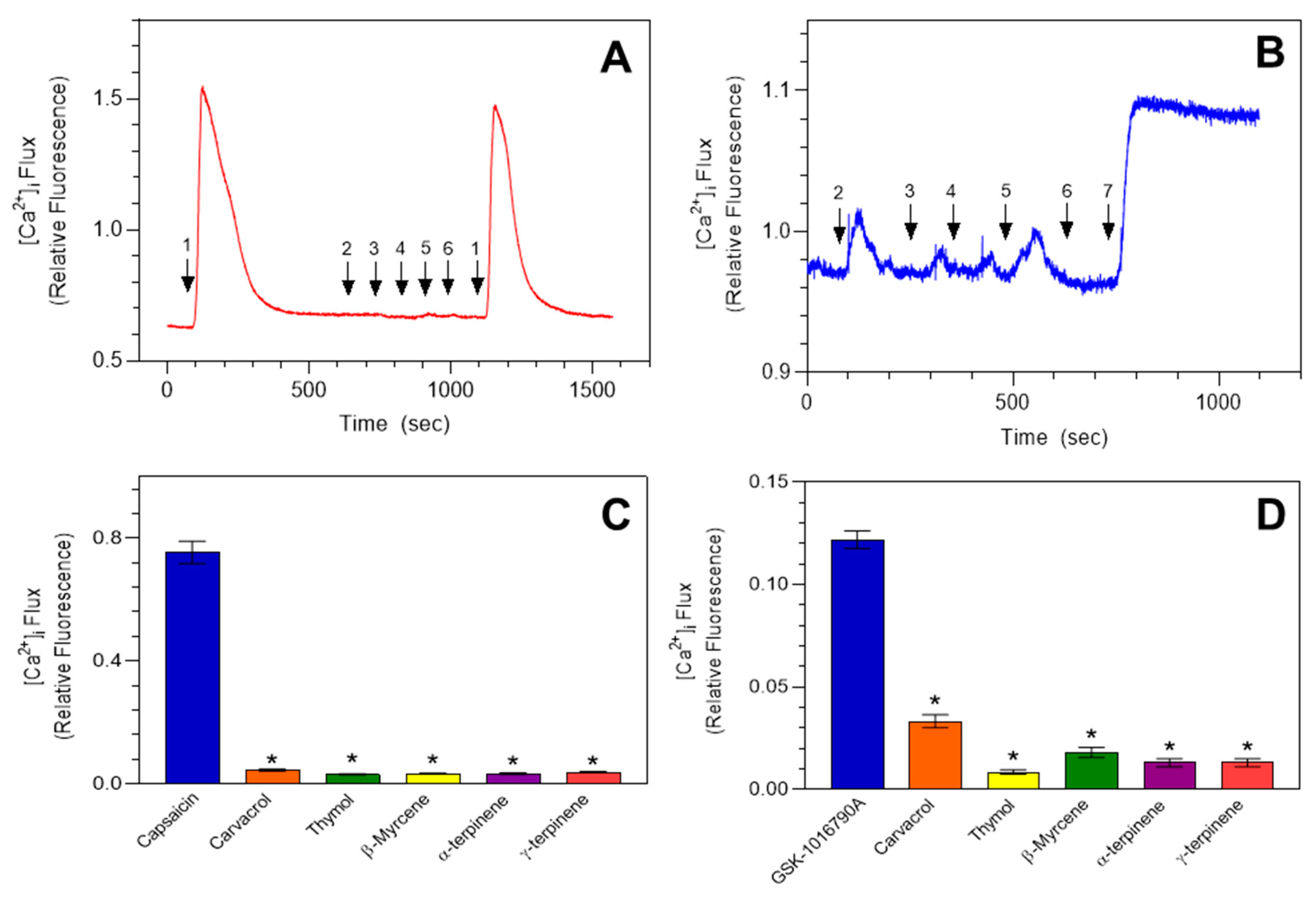
2.3. Effects of Monarda Essential Oil Constituents on TRPA1 Activation
2.4. Molecular Modeling
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chemical Reagents
4.3. Essential Oil Extraction
4.4. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
4.5. Heterologous Expression of TRP Channels in HEK 293 Cells
4.6. “Real-Time” [Ca2+]i Measurements
4.7. Molecular Docking
4.8. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Moerman, D.E. Native American Ethnobotany; Timber Press: Portland, OR, USA, 1998. [Google Scholar]
- Harley, R.M.; Atkins, S.; Budantsev, A.L.; Cantino, P.D.; Conn, B.J.; Grayer, R.; Harley, M.M.; De Kok, R.d.; Krestovskaja, T.d.; Morales, R. Labiatae. In Flowering Plants Dicotyledons; Springer: Berlin/Heidelberg, Germany, 2004; pp. 167–275. [Google Scholar]
- Frezza, C.; Venditti, A.; Serafini, M.; Bianco, A. Phytochemistry, chemotaxonomy, ethnopharmacology, and nutraceutics of Lamiaceae. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2019; Volume 62, pp. 125–178. [Google Scholar]
- Shanaida, M.; Jasicka-Misiak, I.; Makowicz, E.; Stanek, N.; Shanaida, V.; Wieczorek, P.P. Development of high-performance thin layer chromatography method for identification of phenolic compounds and quantification of rosmarinic acid content in some species of the Lamiaceae family. J. Pharm. Bioallied Sci. 2020, 12, 139. [Google Scholar] [CrossRef] [PubMed]
- Lapina, A.S.; Kurkin, V.A. The study of the flavonoid composition of the herb of Monarda fistulosa leaves. Aspir. Vestn. Povolzhiya 2019, 19, 135–142. [Google Scholar] [CrossRef]
- Shanaida, M.; Golembiovska, O. Identification and component analysis of triterpenoids in Monarda fistulosa L. and Ocimum americanum L. (Lamiaceae) aerial parts. Sci.Rise Pharm. Sci. 2018, 26–31. [Google Scholar] [CrossRef]
- Yamada, K.; Murata, T.; Kobayashi, K.; Miyase, T.; Yoshizaki, F. A lipase inhibitor monoterpene and monoterpene glycosides from Monarda punctata. Phytochemistry 2010, 71, 1884–1891. [Google Scholar] [CrossRef]
- Marchioni, I.; Najar, B.; Ruffoni, B.; Copetta, A.; Pistelli, L.; Pistelli, L. Bioactive Compounds and Aroma Profile of Some Lamiaceae Edible Flowers. Plants 2020, 9, 691. [Google Scholar] [CrossRef]
- Kondi, T.; Nakane, Y.; Tamura, H.; Goto, T.; Eugster, C. Structure of monardaein, a bis-malonylated anthocyanin isolated from golden balm, Monarda didyma. Tetrahedron Lett. 1985, 26, 5879–5882. [Google Scholar] [CrossRef]
- Mattarelli, P.; Epifano, F.; Minardi, P.; Di Vito, M.; Modesto, M.; Barbanti, L.; Bellardi, M.G. Chemical composition and antimicrobial activity of essential oils from aerial parts of Monarda didyma and Monarda fistulosa cultivated in Italy. J. Essent. Oil Bear. Plants 2017, 20, 76–86. [Google Scholar] [CrossRef]
- Laquale, S.; Avato, P.; Argentieri, M.; Bellardi, M.; D’Addabbo, T. Nematotoxic activity of essential oils from Monarda species. J. Pest Sci. 2018, 91, 1115–1125. [Google Scholar] [CrossRef]
- Mickienė, R.; Bakutis, B.; Maruška, A.; Ragažinskienė, O.; Kaškonienė, V. Activity of the volatile secondary metabolites of Monarda didyma L. and Myrhis odorata L. plants against aspergillus species micromycetes. In Proceedings of the Modern Aspects And Applications of Chromatography in Education, Research Industry: I International Conference, Daugavpils, Latvia, 16–18 October 2013; Daugavpils University: Daugavpils, Latvia, 2013. [Google Scholar]
- Adebayo, O.; Bélanger, A.; Khanizadeh, S. Variable inhibitory activities of essential oils of three Monarda species on the growth of Botrytis cinerea. Can. J. Plant Sci. 2013, 93, 987–995. [Google Scholar] [CrossRef]
- Ricci, D.; Epifano, F.; Fraternale, D. The Essential Oil of Monarda didyma L.(Lamiaceae) Exerts Phytotoxic Activity in Vitro against Various Weed Seed. Molecules 2017, 22, 222. [Google Scholar] [CrossRef] [Green Version]
- Zhilyakova, E.; Novikov, O.; Naumenko, E.; Krichkovskaya, L.; Kiseleva, T.; Timoshenko, E.Y.; Novikova, M.Y.; Litvinov, S. Study of Monarda fistulosa essential oil as a prospective antiseborrheic agent. Bull. Exp. Biol. Med. 2009, 612–614. [Google Scholar] [CrossRef]
- Tabanca, N.; Bernier, U.R.; Ali, A.; Wang, M.; Demirci, B.; Blythe, E.K.; Khan, S.I.; Baser, K.H.C.; Khan, I.A. Bioassay-guided investigation of two Monarda essential oils as repellents of yellow fever mosquito Aedes aegypti. J. Agric. Food Chem. 2013, 61, 8573–8580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casian, I.; Casian, A.; Valica, V. Obtaining the Wild bergamot essential oil with high content of thymoquinone. Mold. Med. J. 2020, 63, 40–43. [Google Scholar] [CrossRef]
- Colţun, M.; Gille, E.; Necula, R.; Grigoraș, V. Biological Study And Chemical Composition of Monarda fistulosa L. essential oil. Studia Univ. Mold.-Științe R. Și Ale Nat. 2018, 6, 35–38. [Google Scholar]
- Shanaida, M. Antioxidant activity of essential oils obtained from aerial part of some Lamiaceae species. Int. J. Green Pharm. (IJGP) 2018, 12, 200–204. [Google Scholar] [CrossRef]
- Mazza, G.; Kiehn, F.; Marshall, H. Monarda: A source of geraniol, linalool, thymol and carvacrol-rich essential oils. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: Hoboken, NY, USA, 1993; pp. 628–631. [Google Scholar]
- Li, H.; Yang, T.; Li, F.-Y.; Yao, Y.; Sun, Z.-M. Antibacterial activity and mechanism of action of Monarda punctata essential oil and its main components against common bacterial pathogens in respiratory tract. Int. J. Clin. Exp. Pathol. 2014, 7, 7389–7398. [Google Scholar]
- Shen, Q.; Zhou, W.; Li, H.; Hu, L.; Mo, H. ROS involves the fungicidal actions of thymol against spores of Aspergillus flavus via the induction of nitric oxide. PLoS ONE 2016, 11, e0155647. [Google Scholar] [CrossRef]
- Di Vito, M.; Bellardi, M.G.; Mondello, F.; Modesto, M.; Michelozzi, M.; Bugli, F.; Sanguinetti, M.; Sclocchi, M.C.; Sebastiani, M.L.; Biffi, S. Monarda citriodora hydrolate vs essential oil comparison in several anti-microbial applications. Ind. Crop. Prod. 2019, 128, 206–212. [Google Scholar] [CrossRef]
- Lu, Z.G.; Li, X.H.; Li, W. Chemical composition of antibacterial activity of essential oil from Monarda citriodora flowers. In Advanced Materials Research; Trans Tech Publications Ltd: Zurich, Switzerland, 2011; Volume 183, pp. 920–923. [Google Scholar]
- Singh, A.; Chaudhari, A.K.; Das, S.; Dubey, N.K. Nanoencapsulated Monarda citriodora Cerv. ex Lag. essential oil as potential antifungal and antiaflatoxigenic agent against deterioration of stored functional foods. J. Food Sci. Technol. 2020, 57, 2863–2876. [Google Scholar] [CrossRef]
- Pathania, A.S.; Guru, S.K.; Verma, M.; Sharma, C.; Abdullah, S.T.; Malik, F.; Chandra, S.; Katoch, M.; Bhushan, S. Disruption of the PI3K/AKT/mTOR signaling cascade and induction of apoptosis in HL-60 cells by an essential oil from Monarda citriodora. Food Chem. Toxicol. 2013, 62, 246–254. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Deans, S.G. Chemical Composition, Antimicrobial and In Vitro Antioxidant Properties of Monarda citriodora var. citriodora, Myristica fragrans, Origanum vulgare ssp. hirtum, Pelargonium sp. and Thymus zygis Oils. J. Essent. Oil Res. 2004, 16, 145–150. [Google Scholar] [CrossRef]
- Fujisaki, R.; Kamei, K.; Yamamura, M.; Nishiya, H.; Inouye, S.; Takahashi, M.; Abe, S. In vitro and in vivo anti-plasmodial activity of essential oils, including hinokitiol. Southeast Asian J. Trop. Med. Public. Health 2012, 43, 270–279. [Google Scholar] [PubMed]
- Prakash, V. Spices: Leafy Spices; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef] [Green Version]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Koyama, S.; Purk, A.; Kaur, M.; Soini, H.A.; Novotny, M.V.; Davis, K.; Kao, C.C.; Matsunami, H.; Mescher, A. Beta-caryophyllene enhances wound healing through multiple routes. PLoS ONE 2019, 14, e0216104. [Google Scholar] [CrossRef] [Green Version]
- Simões, R.R.; dos Santos Coelho, I.; Junqueira, S.C.; Pigatto, G.R.; Salvador, M.J.; Santos, A.R.S.; de Faria, F.M. Oral treatment with essential oil of Hyptis spicigera Lam.(Lamiaceae) reduces acute pain and inflammation in mice: Potential interactions with transient receptor potential (TRP) ion channels. J. Ethnopharmacol. 2017, 200, 8–15. [Google Scholar] [CrossRef]
- Mancuso, G.; Borgonovo, G.; Scaglioni, L.; Bassoli, A. Phytochemicals from Ruta graveolens activate TAS2R bitter taste receptors and TRP channels involved in gustation and nociception. Molecules 2015, 20, 18907–18922. [Google Scholar] [CrossRef] [Green Version]
- Earley, S.; Gonzales, A.L.; Garcia, Z.I. A dietary agonist of transient receptor potential cation channel V3 elicits endothelium-dependent vasodilation. Mol. Pharmacol. 2010, 77, 612–620. [Google Scholar] [CrossRef] [Green Version]
- Baraldi, P.G.; Preti, D.; Materazzi, S.; Geppetti, P. Transient receptor potential ankyrin 1 (TRPA1) channel as emerging target for novel analgesics and anti-inflammatory agents. J. Med. Chem. 2010, 53, 5085–5107. [Google Scholar] [CrossRef] [PubMed]
- Perry, N.B.; Anderson, R.E.; Brennan, N.J.; Douglas, M.H.; Heaney, A.J.; McGimpsey, J.A.; Smallfield, B.M. Essential oils from dalmatian sage (Salvia officinalis L.): Variations among individuals, plant parts, seasons, and sites. J. Agric. Food Chem. 1999, 47, 2048–2054. [Google Scholar] [CrossRef]
- Özek, G.; Schepetkin, I.A.; Utegenova, G.A.; Kirpotina, L.N.; Andrei, S.R.; Özek, T.; Başer, K.H.C.; Abidkulova, K.T.; Kushnarenko, S.V.; Khlebnikov, A.I.; et al. Chemical composition and phagocyte immunomodulatory activity of Ferula iliensis essential oils. J. Leukoc. Biol. 2017, 101, 1361–1371. [Google Scholar] [CrossRef] [Green Version]
- Schepetkin, I.A.; Özek, G.; Özek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Chemical Composition and Immunomodulatory Activity of Hypericum perforatum Essential Oils. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Zamureenko, V.; Klyuev, N.; Bocharov, B.; Kabanov, V.; Zakharov, A. An investigation of the component composition of the essential oil of Monarda fistulosa. Chem. Nat. Compd. 1989, 25, 549–551. [Google Scholar] [CrossRef]
- Taborsky, J.; Kunt, M.; Kloucek, P.; Lachman, J.; Zeleny, V.; Kokoska, L. Identification of potential sources of thymoquinone and related compounds in Asteraceae, Cupressaceae, Lamiaceae, and Ranunculaceae families. Open Chem. 2012, 10, 1899–1906. [Google Scholar] [CrossRef] [Green Version]
- Rohlfsen, W.G. Method for Cultivation of Monarda fistulosa for Production of Thymoquinone. Google Patents US9073824B2, 26 January 2012. [Google Scholar]
- Pang, J.; Shen, N.; Yan, F.; Zhao, N.; Dou, L.; Wu, L.-C.; Seiler, C.L.; Yu, L.; Yang, K.; Bachanova, V. Thymoquinone exerts potent growth-suppressive activity on leukemia through DNA hypermethylation reversal in leukemia cells. Oncotarget 2017, 8, 34453. [Google Scholar] [CrossRef] [Green Version]
- Farkhondeh, T.; Samarghandian, S.; Borji, A. An overview on cardioprotective and anti-diabetic effects of thymoquinone. Asian Pac. J. Trop. Med. 2017, 10, 849–854. [Google Scholar] [CrossRef]
- Sovova, H.; Sajfrtova, M.; Topiar, M. Supercritical CO2 extraction of volatile thymoquinone from Monarda didyma and M. fistulosa herbs. J. Supercrit. Fluids 2015, 105, 29–34. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Raisinghani, M. Nociceptors in cardiovascular functions: Complex interplay as a result of cyclooxygenase inhibition. Mol. Pain 2006, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Safdar, M.; Khan, M.M.A.; Khattak, K.N.; Anderson, R.A. Cinnamon improves glucose and lipids of people with type 2 diabetes. Diabetes Care 2003, 26, 3215–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hlebowicz, J.; Darwiche, G.; Björgell, O.; Almér, L.-O. Effect of cinnamon on postprandial blood glucose, gastric emptying, and satiety in healthy subjects. Am. J. Clin. Nutr. 2007, 85, 1552–1556. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.-S.; Zhong, L.; Hsieh, T.-H.; Abooj, M.; Bishnoi, M.; Hughes, L.; Premkumar, L.S. Expression of transient receptor potential ankyrin 1 (TRPA1) and its role in insulin release from rat pancreatic beta cells. PLoS ONE 2012, 7, e38005. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Bishnoi, M. Disease-related changes in TRPV1 expression and its implications for drug development. Curr. Top. Med. Chem. 2011, 11, 2192–2209. [Google Scholar] [CrossRef]
- Pabbidi, R.M.; Yu, S.-Q.; Peng, S.; Khardori, R.; Pauza, M.E.; Premkumar, L.S. Influence of TRPV1 on diabetes-induced alterations in thermal pain sensitivity. Mol. Pain 2008, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Van Buren, J.J.; Bhat, S.; Rotello, R.; Pauza, M.E.; Premkumar, L.S. Sensitization and translocation of TRPV1 by insulin and IGF-I. Mol. Pain 2005, 1, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Materazzi, S.; Fusi, C.; Benemei, S.; Pedretti, P.; Patacchini, R.; Nilius, B.; Prenen, J.; Creminon, C.; Geppetti, P.; Nassini, R. TRPA1 and TRPV4 mediate paclitaxel-induced peripheral neuropathy in mice via a glutathione-sensitive mechanism. Pflügers Arch. Eur. J. Physiol. 2012, 463, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Eid, S.R.; Crown, E.D.; Moore, E.L.; Liang, H.A.; Choong, K.-C.; Dima, S.; Henze, D.A.; Kane, S.A.; Urban, M.O. HC-030031, a TRPA1 selective antagonist, attenuates inflammatory- and neuropathy-induced mechanical hypersensitivity. Mol. Pain 2008, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Szallasi, A.; Blumberg, P.M. Vanilloid (Capsaicin) Receptors and Mechanisms. Pharmacol. Rev. 1999, 51, 159–212. [Google Scholar]
- Baratchi, S.; Keov, P.; Darby, W.G.; Lai, A.; Khoshmanesh, K.; Thurgood, P.; Vahidi, P.; Ejendal, K.; McIntyre, P. The TRPV4 Agonist GSK1016790A Regulates the Membrane Expression of TRPV4 Channels. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepetkin, I.A.; Kushnarenko, S.V.; Ozek, G.; Kirpotina, L.N.; Utegenova, G.A.; Kotukhov, Y.A.; Danilova, A.N.; Ozek, T.; Baser, K.H.; Quinn, M.T. Inhibition of Human Neutrophil Responses by the Essential Oil of Artemisia kotuchovii and Its Constituents. J Agric Food Chem 2015, 63, 4999–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, C.E.; Armache, J.-P.; Gao, Y.; Cheng, Y.; Julius, D. Structure of the TRPA1 ion channel suggests regulatory mechanisms. Nature 2015, 520, 511–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarenga, E.M.; Souza, L.K.; Araújo, T.S.; Nogueira, K.M.; Sousa, F.B.M.; Araújo, A.R.; Martins, C.S.; Pacífico, D.M.; Brito, G.A.d.C.; Souza, E.P. Carvacrol reduces irinotecan-induced intestinal mucositis through inhibition of inflammation and oxidative damage via TRPA1 receptor activation. Chem. Biol. Interact. 2016, 260, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Grzeszczuk, M.; Wesolowska, A.; Stefaniak, A. Biological value and essential oil composition of two Monarda species flowers. Acta Sci. Pol. Hortorum Cultus 2020, 19, 105–119. [Google Scholar] [CrossRef]
- Duke, J.A. Handbook of Medicinal Herbs; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Mazza, G.; Marshall, H. Geraniol, linalool, thymol and carvacrol-rich essential oils from Monarda hybrids. J. Essent. Oil Res. 1992, 4, 395–400. [Google Scholar] [CrossRef]
- Lawrence, B. Uncommon essential oils as sources of common natural aroma chemicals. Perfum. Flavorist 1985, 10, 45–58. [Google Scholar]
- Premkumar, L.S. Transient receptor potential channels as targets for phytochemicals. ACS Chem. Neurosci. 2014, 5, 1117–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Delling, M.; Jun, J.C.; Clapham, D.E. Oregano, thyme and clove-derived flavors and skin sensitizers activate specific TRP channels. Nat. Neurosci. 2006, 9, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Shabnum, S.; Wagay, M.G. Essential oil composition of Thymus vulgaris L. and their uses. J. Res. Dev. 2011, 11, 83–94. [Google Scholar]
- Varel, V.; Miller, D. Effect of carvacrol and thymol on odor emissions from livestock wastes. Water Sci. Technol. 2001, 44, 143–148. [Google Scholar] [CrossRef]
- Hotta, M.; Nakata, R.; Katsukawa, M.; Hori, K.; Takahashi, S.; Inoue, H. Carvacrol, a component of thyme oil, activates PPARalpha and gamma and suppresses COX-2 expression. J. Lipid Res. 2010, 51, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Parnas, M.; Peters, M.; Dadon, D.; Lev, S.; Vertkin, I.; Slutsky, I.; Minke, B. Carvacrol is a novel inhibitor of Drosophila TRPL and mammalian TRPM7 channels. Cell Calcium 2009, 45, 300–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Mariri, A.; Swied, G.; Oda, A.; Al Hallab, L. Antibacterial activity of thymus syriacus boiss essential oil and its components against some Syrian gram-negative bacteria isolates. Iran. J. Med. Sci. 2013, 38, 180. [Google Scholar]
- Govindarajan, M.; Sivakumar, R.; Rajeswary, M.; Veerakumar, K. Mosquito larvicidal activity of thymol from essential oil of Coleus aromaticus Benth. against Culex tritaeniorhynchus, Aedes albopictus, and Anopheles subpictus (Diptera: Culicidae). Parasitol. Res. 2013, 112, 3713–3721. [Google Scholar] [CrossRef]
- Andrade, L.N.; De Sousa, D.P. A review on anti-inflammatory activity of monoterpenes. Molecules 2013, 18, 1227–1254. [Google Scholar] [CrossRef]
- Abdel-Rahman, F.H.; Alaniz, N.M.; Saleh, M.A. Nematicidal activity of terpenoids. J. Environ. Sci. HealthPart B 2013, 48, 16–22. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Monteiro, C.M.; Daemon, E.; Silva, A.M.R.; Maturano, R.; Amaral, C. Acaricide and ovicide activities of thymol on engorged females and eggs of Rhipicephalus (Boophilus) microplus (Acari: Ixodidae). Parasitol. Res. 2010, 106, 615–619. [Google Scholar] [CrossRef]
- Lee, K.G.; Shibamoto, T. Antioxidant activities of volatile components isolated from Eucalyptus species. J. Sci. Food Agric. 2001, 81, 1573–1579. [Google Scholar] [CrossRef]
- Xu, Z.-H.; Wang, C.; Fujita, T.; Jiang, C.-Y.; Kumamoto, E. Action of thymol on spontaneous excitatory transmission in adult rat spinal substantia gelatinosa neurons. Neurosci. Lett. 2015, 606, 94–99. [Google Scholar] [CrossRef]
- Lee, S.; Buber, M.; Yang, Q.; Cerne, R.; Cortes, R.; Sprous, D.; Bryant, R. Thymol and related alkyl phenols activate the hTRPA1 channel. Br. J. Pharmacol. 2008, 153, 1739–1749. [Google Scholar] [CrossRef] [Green Version]
- Karashima, Y.; Damann, N.; Prenen, J.; Talavera, K.; Segal, A.; Voets, T.; Nilius, B. Bimodal action of menthol on the transient receptor potential channel TRPA1. J. Neurosci. 2007, 27, 9874–9884. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Dubin, A.E.; Bursulaya, B.; Viswanath, V.; Jegla, T.J.; Patapoutian, A. Identification of transmembrane domain 5 as a critical molecular determinant of menthol sensitivity in mammalian TRPA1 channels. J. Neurosci. 2008, 28, 9640–9651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upton, R.; Craker, L.; ElSohly, M.; Romm, A.; Russo, E.; Sexton, M. Cannabis inflorescence: Standards of identity, analysis, and quality control. In American Herbal Pharmacopoeia and Therapeutic Compendium; American Herbal Pharmacopoeia: Scotts Valley, CA, USA, 2014; pp. 1–15. [Google Scholar]
- Hartsel, J.A.; Eades, J.; Hickory, B.; Makriyannis, A. Cannabis sativa and Hemp. In Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2016; pp. 735–754. [Google Scholar]
- Heblinski, M.; Santiago, M.; Fletcher, C.; Stuart, J.; Connor, M.; McGregor, I.S.; Arnold, J.C. Terpenoids Commonly Found in Cannabis sativa Do Not Modulate the Actions of Phytocannabinoids or Endocannabinoids on TRPA1 and TRPV1 Channels. Cannabis Cannabinoid Res. 2020. [Google Scholar] [CrossRef] [Green Version]
- Jansen, C.; Shimoda, L.; Kawakami, J.; Ang, L.; Bacani, A.; Baker, J.; Badowski, C.; Speck, M.; Stokes, A.; Small-Howard, A. Myrcene and terpene regulation of TRPV1. Channels 2019, 13, 344–366. [Google Scholar] [CrossRef] [Green Version]
- Schepetkin, I.A.; Kushnarenko, S.V.; Özek, G.; Kirpotina, L.N.; Sinharoy, P.; Utegenova, G.A.; Abidkulova, K.T.; Özek, T.; Başer, K.H.s.C.; Kovrizhina, A.R. Modulation of human neutrophil responses by the essential oils from Ferula akitschkensis and their constituents. J. Agric. Food Chem. 2016, 64, 7156–7170. [Google Scholar] [CrossRef] [Green Version]
- Cortright, D.N.; Szallasi, A. Biochemical pharmacology of the vanilloid receptor TRPV1. Eur. J. Biochem. 2004, 271, 1814–1819. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.; Wyse, B.D.; Roberts-Thomson, S.J.; Monteith, G.R.; Cabot, P.J. Mechanisms involved in potentiation of transient receptor potential vanilloid 1 responses by ethanol. Eur. J. Pain 2008, 12, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Gholamnezhad, Z.; Havakhah, S.; Boskabady, M.H. Preclinical and clinical effects of Nigella sativa and its constituent, thymoquinone: A review. J. Ethnopharmacol. 2016, 190, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Mihara, S.; Shibamoto, T. The role of flavor and fragrance chemicals in TRPA1 (transient receptor potential cation channel, member A1) activity associated with allergies. Allergy Asthma Clin. Immunol. 2015, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Özek, G.; Ishmuratova, M.; Tabanca, N.; Radwan, M.M.; Göger, F.; Özek, T.; Wedge, D.E.; Becnel, J.J.; Cutler, S.J.; Can Başer, K.H. One-step multiple component isolation from the oil of Crinitaria tatarica (Less.) S ojak by preparative capillary gas chromatography with characterization by spectroscopic and spectrometric techniques and evaluation of biological activity. J. Sep. Sci. 2012, 35, 650–660. [Google Scholar] [CrossRef]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monarda spp. | Biological Activity | Major Constituents | Ref. |
|---|---|---|---|
| M. didyma L. | Nematotoxic | Carvacrol, γ-terpinene, o-cymene, and thymol | [11] |
| Antifungal | Thymol, carvacrol, α-terpinene, p-cymene, γ-terpinene | [12,13] | |
| Phytotoxic | Essential oil | [14] | |
| Antimicrobial | Thymol | [10] | |
| - | Thymol, thymol methyl ether | [8] | |
| M. fistulosa L. | Nematotoxic | Carvacrol, γ-terpinene, o-cymene, and thymol | [11] |
| Antiseborrheic | Essential oil | [15] | |
| Repellent | Carvacrol, thymol, eugenol, carvacrol methyl ether | [16] | |
| - | Thymol, carvacrol, thymoquinone | [17,18] | |
| Antifungal | Geraniol, geranial, geranyl formate, p-cymene | [13] | |
| Antioxidant | Essential oil, thymol, carvacrol | [19] | |
| Antimicrobial | Thymol | [10] | |
| M. fistulosa, M. didyma | - | Geraniol, linalool, thymol, carvacrol, 1,8-cineole | [20] |
| M. bradburiana Beck | Repellent | Carvacrol, carvacrol methyl ether, thymol, eugenol | [16] |
| M. punctata L. | Antilipase | Carvacrol | [7] |
| Antibacterial | Thymol, p-cymene, limonene, carvacrol | [24] | |
| Fungicidal | Thymol | [22] | |
| M. citriodora Cerv. ex Lag. | Antimicrobial | Essential oil, hydrolate | [23] |
| Antibacterial | Thymol, 1,8-cineole, α-phellandrene, p-cymene | [24] | |
| Antifungal | β-Caryophyllene, citral, limonene, cis-verbenol | [25] | |
| Anticancer (in vitro) | Essential oil, thymol | [26] | |
| M. citriodora var. citriodora Cerv. ex Lag. | Antioxidant, antimicrobial | Essential oil | [27] |
| M. menthifolia Graham | - | p-Cymene, carvacrol, hydrothymoquinone, geraniol, linalool | [29] |
| M. pectinata Nutt. | - | Carvacrol | [29] |
| Monarda spp. | Antiplasmodial | Essential oil, thymoquinone | [28] |
| # | RRI | Compound | MEOFl | MEOLv | # | RRI | Compound | MEOFl | MEOLv |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 1032 | α-Pinene | 0.7 | 0.4 | 23 | 1497 | α-Copaene | T | t |
| 2 | 1035 | α-Thujene | 2.4 | 1.3 | 24 | 1535 | β-Bourbonene | T | 0.1 |
| 3 | 1076 | Camphene | 0.1 | 0.1 | 25 | 1553 | Linalool | 0.1 | 0.1 |
| 4 | 1118 | β-Pinene | 0.2 | 0.1 | 26 | 1556 | cis-Sabinene hydrate | 0.2 | 0.2 |
| 5 | 1132 | Sabinene | 0.3 | 0.1 | 27 | 1571 | trans-p-Menth-2-en-1-ol | 0.1 | t |
| 6 | 1159 | δ-3-Carene | 0.2 | 0.1 | 28 | 1590 | Bornyl acetate | T | |
| 7 | 1167 | 3-Heptanone | t | 29 | 1611 | Terpinen-4-ol | 1.1 | ||
| 8 | 1174 | β-Myrcene | 1.4 | 0.8 | 30 | 1614 | Methyl carvacrol | 1.1 | 1.4 |
| 9 | 1176 | α-Phellandrene | 0.3 | 0.2 | 31 | 1638 | cis-p-Menth-2-en-1-ol | T | |
| 10 | 1183 | p-Mentha-1,7(8)-diene | t | 32 | 1689 | trans-Piperitol | 0.2 | ||
| 11 | 1188 | α-Terpinene | 3.8 | 1.7 | 33 | 1704 | γ-Muurolene | t | |
| 12 | 1203 | Limonene | 0.9 | 0.5 | 34 | 1706 | α-Terpineol | 0.2 | 0.1 |
| 13 | 1218 | β-Phellandrene | 0.3 | 0.2 | 35 | 1719 | Borneol | 0.2 | 0.1 |
| 14 | 1255 | γ-Terpinene | 6.8 | 2.5 | 36 | 1726 | Germacrene D | 0.3 | 0.2 |
| 15 | 1266 | 3-Octanone | t | 37 | 1773 | δ-Cadinene | t | ||
| 16 | 1280 | p-Cymene | 25.6 | 13.1 | 38 | 1802 | Cumin aldehyde | t | |
| 17 | 1290 | Terpinolene | 0.1 | 39 | 1864 | p-Cymen-8-ol | 0.1 | ||
| 18 | 1393 | 3-Octanol | 0.1 | 40 | 1889 | Ascaridole | t | ||
| 19 | 1400 | Nonanal | t | 41 | 1940 | 4-Isopropyl salicylaldehyde | t | ||
| 20 | 1408 | 1,3,8-p-Menthatriene | t | 42 | 1969 | cis-Jasmone | 0.1 | ||
| 21 | 1452 | 1-Octen-3-ol | 1.5 | 0.9 | 43 | 2113 | Cumin alcohol | t | |
| 22 | 1474 | trans-Sabinene hydrate | 1 | 0.8 | 44 | 2198 | Thymol | 3.1 | 3.3 |
| 45 | 2239 | Carvacrol | 45.7 | 71.5 |
| Total | MEOFl | MEOLv |
|---|---|---|
| % | ||
| 99.8 | 100 | |
| Monoterpene hydrocarbons | 43.1 | 21.1 |
| Oxygenated monoterpenes | 54.8 | 77.7 |
| Sesquiterpene hydrocarbons | 0.3 | 0.3 |
| Miscellaneous compounds | 1.6 | 0.9 |
| Compound | Composition | HEK Cells | ||
|---|---|---|---|---|
| MEOFl | MEOLv | rTRPA1 | NT | |
| % | [Ca2+]i Change | |||
| α-Pinene | 0.7 | 0.4 | 0.054 ± 0.014 | 0.013 ± 0.002 |
| Camphene | 0.1 | 0.1 | 0.014 ± 0.004 | 0.009 ± 0.001 |
| β-Pinene | 0.2 | 0.1 | 0.012 ± 0.002 | 0.011 ± 0.001 |
| Sabinene | 0.3 | 0.1 | 0.029 ± 0.005 | 0.034 ± 0.004 |
| β-Myrcene | 1.4 | 0.8 | 0.111 ± 0.003 | 0.005 ± 0.001 |
| α-Terpinene | 3.8 | 1.7 | 0.007 ± 0.001 | 0.006 ± 0.001 |
| (-) Limonene | 0.9 | 0.5 | 0.008 ± 0.002 | 0.007 ± 0.001 |
| (+) Limonene | - | - | 0.011 ± 0.002 | 0.009 ± 0.002 |
| γ-Terpinene | 6.8 | 2.5 | 0.034 ± 0.005 | 0.014 ± 0.001 |
| p-Cymene | 25.6 | 13.1 | 0.071 ± 0.011 | 0.027 ± 0.001 |
| Terpinolene | 0.1 | - | 0.034 ± 0.007 | 0.022 ± 0.003 |
| Linalool | 0.1 | 0.1 | 0.037 ± 0.007 | 0.002 ± 0.0003 |
| Terpinen-4-ol | 1.1 | - | 0.046 ± 0.007 | 0.013 ± 0.002 |
| α-Terpineol | 0.2 | 0.1 | 0.010 ± 0.002 | 0.004 ± 0.001 |
| Thymol | 3.1 | 3.3 | 0.849 ± 0.013 | 0.026 ± 0.002 |
| Carvacrol | 45.7 | 71.5 | 0.340 ± 0.012 | 0.061 ± 0.018 |
Sample Availability: Samples of the compounds are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, M.; Schepetkin, I.A.; Özek, G.; Özek, T.; Khlebnikov, A.I.; Damron, D.S.; Quinn, M.T. Essential Oils from Monarda fistulosa: Chemical Composition and Activation of Transient Receptor Potential A1 (TRPA1) Channels. Molecules 2020, 25, 4873. https://doi.org/10.3390/molecules25214873
Ghosh M, Schepetkin IA, Özek G, Özek T, Khlebnikov AI, Damron DS, Quinn MT. Essential Oils from Monarda fistulosa: Chemical Composition and Activation of Transient Receptor Potential A1 (TRPA1) Channels. Molecules. 2020; 25(21):4873. https://doi.org/10.3390/molecules25214873
Chicago/Turabian StyleGhosh, Monica, Igor A. Schepetkin, Gulmira Özek, Temel Özek, Andrei I. Khlebnikov, Derek S. Damron, and Mark T. Quinn. 2020. "Essential Oils from Monarda fistulosa: Chemical Composition and Activation of Transient Receptor Potential A1 (TRPA1) Channels" Molecules 25, no. 21: 4873. https://doi.org/10.3390/molecules25214873