
Fragaria × ananassa cv. Senga Sengana Leaf: An Agricultural Waste with Antiglycation Potential and High Content of Ellagitannins, Flavonols, and 2-Pyrone-4,6-dicarboxylic Acid


Abstract
:
1. Introduction
2. Results
2.1. Hydrolyzable Tannins
2.2. Proanthocyanidins and Flavan-3-Ols
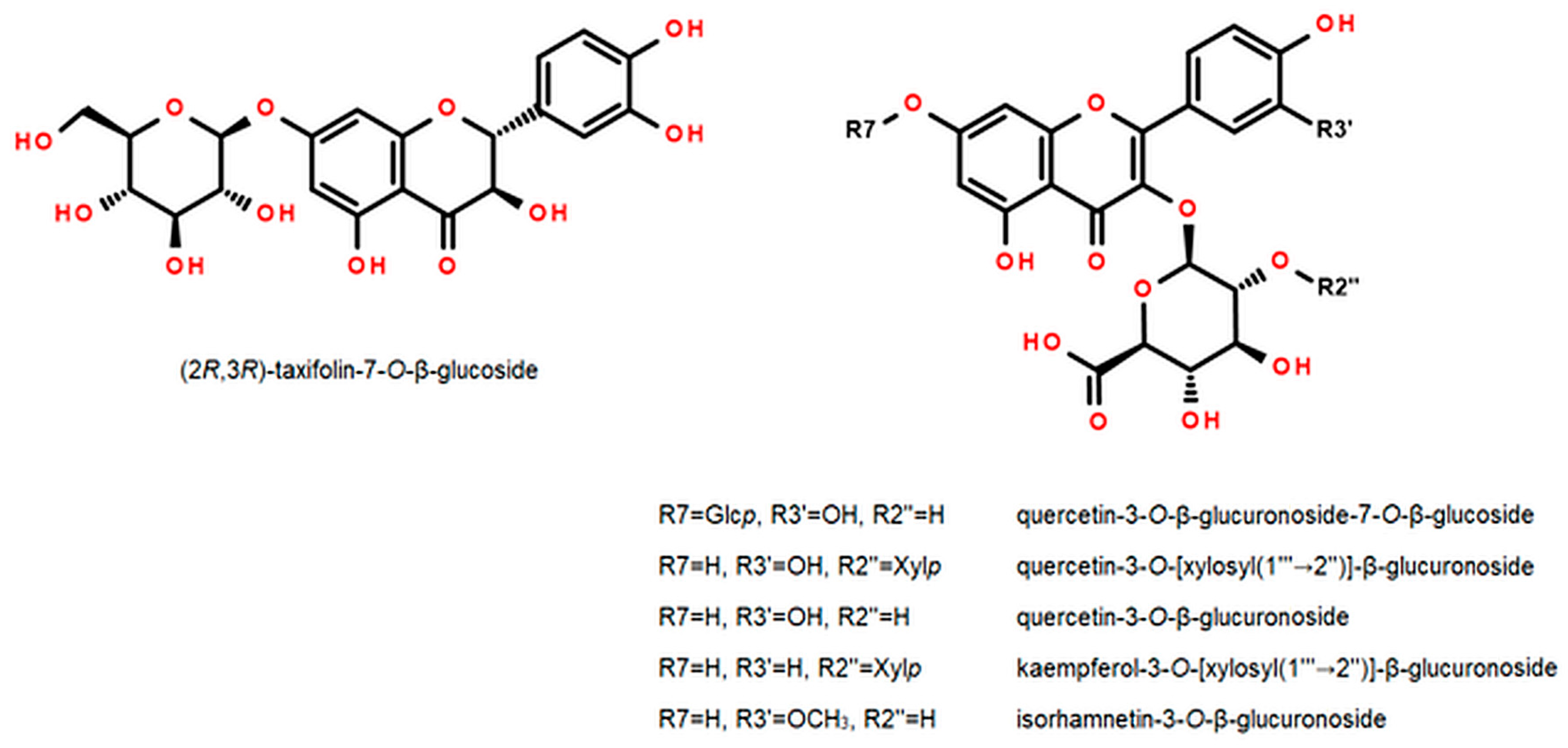
2.3. Flavonoids
2.4. Phenolic and Carboxylic Acids
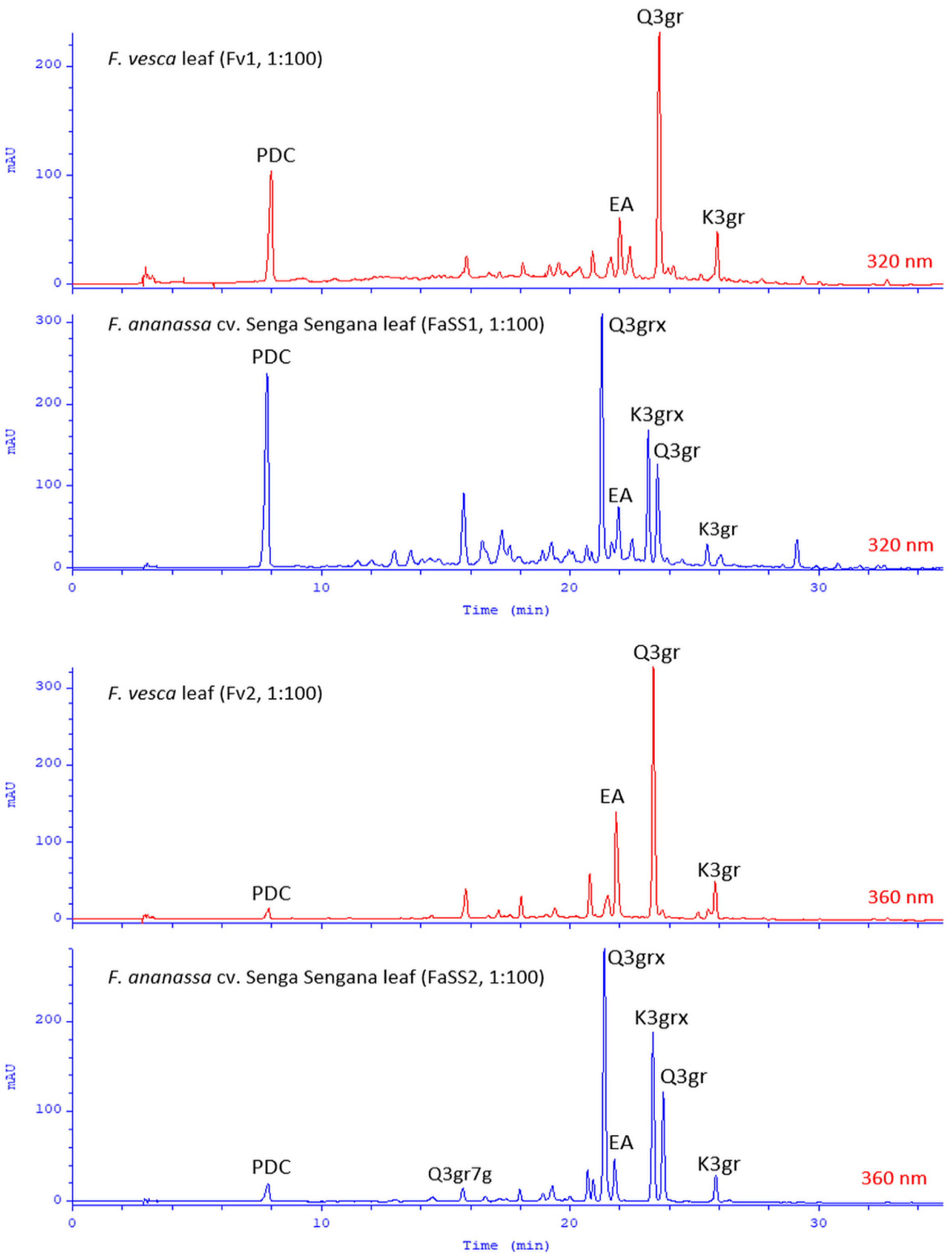
2.5. Chemical Similarity of the Leaves of F. ananassa cv. Senga Sengana and F. vesca
2.6. Antiglycative and Anti-MGO Effects of Flavonols
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Standards
4.3. Plant Material
4.4. Sample Preparation for LC
4.5. Isolation of F. ananassa cv. Senga Sengana Leaf Constituents
4.6. Identification and Quantification
4.6.1. Structure Elucidation Equipment
4.6.2. Chromatography
4.6.3. Acid Hydrolysis
4.6.4. Alkaline Hydrolysis
4.7. Inhibition of Glycation and MGO Trapping In Vitro
4.7.1. Antiglycation Assay in BSA-MGO Model
4.7.2. MGO Trapping Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Appendix A. Details on Isolation of F. ananassa cv. Senga Sengana Leaf Constituents
Appendix B. Details on Identification of F. ananassa cv. Senga Sengana Leaf Constituents
Appendix B.1. Hydrolyzable Tannins
Appendix B.2. Proanthocyanidins and Flavan-3-Ols
Appendix B.3. Flavonoids
Appendix B.4. Phenolic and Carboxylic Acids
References
- Koch, W. Dietary polyphenols—Important non-nutrients in the prevention of chronic noncommunicable diseases. A systematic review. Nutrients 2019, 11, 1039. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and oxidative stress in human diseases: From molecular mechanisms to novel treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef] [PubMed]
- Mortera, R.R.; Bains, Y.; Gugliucci, A. Fructose at the crossroads of the metabolic syndrome and obesity epidemics. Front. Biosci. Landmark 2019, 24, 186–211. [Google Scholar]
- Yang, C.S.; Wang, H.; Sheridan, Z.P. Studies on prevention of obesity, metabolic syndrome, diabetes, cardiovascular diseases and cancer by tea. J. Food Drug Anal. 2018, 26, 1–13. [Google Scholar] [CrossRef]
- Liu, J.; He, Z.; Ma, N.; Chen, Z.Y. Beneficial effects of dietary polyphenols on high-fat diet-induced obesity linking with modulation of gut microbiota. J. Agric. Food Chem. 2020, 68, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Catalkaya, G.; Venema, K.; Lucini, L.; Rocchetti, G.; Delmas, D.; Daglia, M.; De Filippis, A.; Xiao, H.; Quiles, J.L.; Xiao, J.; et al. Interaction of dietary polyphenols and gut microbiota: Microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 2020, 1, 109–133. [Google Scholar] [CrossRef]
- García-Villalba, R.; Vissenaekens, H.; Pitart, J.; Romo-Vaquero, M.; Espín, J.C.; Grootaert, C.; Selma, M.V.; Raes, K.; Smagghe, G.; Possemiers, S.; et al. Gastrointestinal simulation model TWIN-SHIME shows differences between human urolithin-metabotypes in gut microbiota composition, pomegranate polyphenol metabolism, and transport along the intestinal tract. J. Agric. Food Chem. 2017, 65, 5480–5493. [Google Scholar] [CrossRef]
- Widy-Tyszkiewicz, E. Assessment Report on Fragaria Vesca L., Fragaria Moschata Weston, Fragaria Viridis Weston and Fragaria × Ananassa (Weston) Duchesne ex Rozier, Folium. 2018. Available online: https://www.ema.europa.eu/en/documents/herbal-report/final-assessment-report-fragaria-vesca-l-fragaria-moschata-weston-fragaria-viridis-weston-fragaria-x_en.pdf (accessed on 28 July 2022).
- Blaschek, W.; Ebel, S.; Hackental, E.; Holzgrabe, U.; Keller, K.; Reichling, J.; Schulz, J. (Eds.) Fragariae folium. In Hagers Handbuch der Drogen und Arzneistoffe; HagerRom; Springer Medizin Verlag: Heidelberg, Germany, 2006; ISBN 978-3540282327. [Google Scholar]
- Wichtl, M.; Blaschek, W. (Eds.) Fragariae folium. In Teedrogen und Phytopharmaka: Ein Handbuch für die Praxis; Wissenschaftliche Verlagsgesellschaft: Stuttgart, Germany, 2016; pp. 262–263. ISBN 978-3804730687. [Google Scholar]
- Fecka, I. Development of chromatographic methods for determination of agrimoniin and related polyphenols in pharmaceutical products. J. AOAC Int. 2009, 92, 410–418. [Google Scholar] [CrossRef]
- Tridge Report: Strawberry Production. Available online: https://www.tridge.com/intelligences/stawberry/production (accessed on 28 July 2022).
- Bednarska, K.; Fecka, I. Potential of vasoprotectives to inhibit non-enzymatic protein glycation, and reactive carbonyl and oxygen species uptake. Int. J. Mol. Sci. 2021, 22, 10026. [Google Scholar] [CrossRef]
- Clifford, M.N.; Stoupi, S.; Kuhnert, N. Profiling and characterization by LC-MSn of the galloylquinic acids of green tea, tara tannin, and tannic acid. J. Agric. Food Chem. 2007, 55, 2797–2807. [Google Scholar] [CrossRef]
- Nishimura, H.; Nonaka, G.I.; Nishioka, I. Seven quinic acid gallates from Quercus stenophylla. Phytochemistry 1984, 23, 2621–2623. [Google Scholar] [CrossRef]
- Feldman, K.S.; Smith, R.S. Ellagitannin chemistry. First total synthesis of the 2,3- and 4,6-coupled ellagitannin pedunculagin. J. Org. Chem. 1996, 61, 2606–2612. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Kuwahara, M.; Memon, M.U.; Shingu, T. Agrimoniin and potentillin, an ellagitannin dimer and monomer having an α-glucose core. J. Chem. Soc. Chem. Commun. 1982, 3, 162–163. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Kuwahara, M.; Memon, M.U.; Shingu, T. Tannins of rosaceous medicinal plants. I. Structures of potentillin, agrimonic acids A and B, and agrimoniin, a dimeric ellagitannin. Chem. Pharm. Bull. (Tokyo) 1984, 32, 2165–2173. [Google Scholar] [CrossRef]
- Sakushima, A.; Ohno, K.; Coskun, M.; Seki, K.I.; Ohkura, K. Separation and identification of taxifolin 3-O-glucoside isomers from Chamaecyparis obtusa (Cupressaceae). Nat. Prod. Lett. 2002, 16, 383–387. [Google Scholar] [CrossRef]
- Felser, C.; Schimmer, O. Flavonoid glycosides from Alchemilla speciosa. Planta Med. 1999, 65, 668–670. [Google Scholar] [CrossRef]
- Abe, T.; Yoshida, S.; Kawabata, T.; Takano, F.; Ohta, T. Study of inhibitory effect of polyphenols from Fragaria ananassa on fat accumulation in rat white adipocyte. JJCAM 2018, 15, 1–7. [Google Scholar] [CrossRef]
- Ossipov, V.; Nurmi, K.; Loponen, J.; Prokopiev, N.; Haukioja, E.; Pihlaja, K. HPLC isolation and identification of flavonoids from white birch Betula pubescens leaves. Biochem. Syst. Ecol. 1995, 23, 213–222. [Google Scholar] [CrossRef]
- Im, S.H.; Wang, Z.; Lim, S.S.; Lee, O.-H.; Kang, I.-J. Bioactivity-guided isolation and identification of anti-adipogenic compounds from Sanguisorba officinalis. Pharm. Biol. 2017, 55, 2057–2064. [Google Scholar] [CrossRef]
- Timmers, M.; Urban, S. On-line (HPLC-NMR) and off-line phytochemical profiling of the Australian plant, Lasiopetalum macrophyllum. Nat. Prod. Commun. 2011, 6, 1605–1616. [Google Scholar] [CrossRef]
- Wilkes, S.; Glasl, H. Isolation, characterization, and systematic significance of 2-pyrone-4,6-dicarboxylic acid in Rosaceae. Phytochemistry 2001, 58, 441–449. [Google Scholar] [CrossRef]
- Ishimaru, K.; Omoto, T.; Asai, I.; Ezaki, K.; Shimomura, K. Taxifolin 3-arabinoside from Fragaria × ananassa. Phytochemistry 1995, 40, 345–347. [Google Scholar] [CrossRef]
- Kårlund, A.; Salminen, J.P.; Koskinen, P.; Ahern, J.R.; Karonen, M.; Tiilikkala, K.; Karjalainen, R.O. Polyphenols in strawberry (Fragaria × ananassa) leaves induced by plant activators. J. Agric. Food Chem. 2014, 62, 4592–4600. [Google Scholar] [CrossRef]
- Hanhineva, K.; Rogachev, I.; Kokko, H.; Mintz-Oron, S.; Venger, I.; Kärenlampi, S.; Aharoni, A. Non-targeted analysis of spatial metabolite composition in strawberry (Fragaria × ananassa) flowers. Phytochemistry 2008, 69, 2463–2481. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T.; Iwasaki, M.; Kubo, M.; Orime, T.; Yoshizaki, M.; Naruhashi, N. Hydrolysable tannins as chemotaxonomic markers in the Rosaceae. Phytochemistry 1992, 31, 3091–3096. [Google Scholar] [CrossRef]
- Josuttis, M.; Verrall, S.; Stewart, D.; Krüger, E.; McDougall, G.J. Genetic and environmental effects on tannin composition in strawberry (Fragaria × ananassa) cultivars grown in different European locations. J. Agric. Food Chem. 2013, 61, 790–800. [Google Scholar] [CrossRef]
- Hatano, T.; Yazaki, K.; Okonogi, A.; Okuda, T. Tannins of Stachyurus species. II. Praecoxins A, B, C and D, four new hydrolyzable tannins from Stachyurus praecox leaves. Chem. Pharm. Bull. (Tokyo) 1991, 39, 1689–1693. [Google Scholar] [CrossRef]
- Karlińska, E.; Pecio, Ł.; Macierzyński, J.; Stochmal, A.; Kosmala, M. Structural elucidation of the ellagitannin with a molecular weight of 2038 isolated from strawberry fruit (Fragaria ananassa Duch.) and named fragariin A. Food Chem. 2019, 296, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Fecka, I.; Kucharska, A.Z.; Kowalczyk, A. Quantification of tannins and related polyphenols in commercial products of tormentil (Potentilla tormentilla). Phytochem. Anal. 2015, 26, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Tanaka, K.; Xin-min, C.; Okuda, T. Dimeric ellagitannins, laevigatins E, F and G, from Rosa laevigata. Phytochemistry 1989, 28, 2451–2454. [Google Scholar] [CrossRef]
- Yoshida, T.; Tanaka, K.; Chen, X.-M.; Okuda, T. Tannis of rosaceous medicinal plants. V. Hydrolyzable tannis with dehydrodigalloyl group from Rosa laevigata Michx. Chem. Pharm. Bull. (Tokyo) 1989, 37, 920–924. [Google Scholar] [CrossRef]
- Aaby, K.; Mazur, S.; Nes, A.; Skrede, G. Phenolic compounds in strawberry (Fragaria × ananassa Duch.) fruits: Composition in 27 cultivars and changes during ripening. Food Chem. 2012, 132, 86–97. [Google Scholar] [CrossRef]
- Yoshida, T.; Jin, Z.-X.; Okuda, T. Hydrolysable tannin oligomers from Rosa davurica. Phytochemistry 1991, 30, 2747–2752. [Google Scholar] [CrossRef]
- Weber, N.; Veberic, R.; Mikulic-Petkovsek, M.; Stampar, F.; Koron, D.; Munda, A.; Jakopic, J. Metabolite accumulation in strawberry (Fragaria × ananassa Duch.) fruits and runners in response to Colletotrichum nymphaeae infection. Physiol. Mol. Plant Pathol. 2015, 92, 119–129. [Google Scholar] [CrossRef]
- Itoh, T.; Ninomiya, M.; Yasuda, M.; Koshikawa, K.; Deyashiki, Y.; Nozawa, Y.; Akao, Y.; Koketsu, M. Inhibitory effects of flavonoids isolated from Fragaria ananassa Duch. on IgE-mediated degranulation in rat basophilic leukemia RBL-2H3. Bioorg. Med. Chem. 2009, 17, 5374–5379. [Google Scholar] [CrossRef]
- Ninomiya, M.; Itoh, T.; Ishikawa, S.; Saiki, M.; Narumiya, K.; Yasuda, M.; Koshikawa, K.; Nozawa, Y.; Koketsu, M. Phenolic constituents isolated from Fragaria ananassa Duch. inhibit antigen-stimulated degranulation through direct inhibition of spleen tyrosine kinase activation. Bioorg. Med. Chem. 2010, 18, 5932–5937. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Tomise, K.; Aburatani, M.; Onuki, H.; Hirorta, H.; Ishiharajima, E.; Ohta, T. Isolation of cytochrome p450 inhibitors from strawberry fruit, Fragaria ananassa. J. Nat. Prod. 2004, 67, 1839–1841. [Google Scholar] [CrossRef]
- Hanhineva, K.; Soininen, P.; Anttonen, M.J.; Kokko, H.; Rogachev, I.; Aharoni, A.; Laatikainen, R.; Kärenlampi, S. NMR and UPLC-qTOF-MS/MS characterisation of novel phenylethanol derivatives of phenylpropanoid glucosides from the leaves of strawberry (Fragaria × ananassa cv. Jonsok). Phytochem. Anal. 2009, 20, 353–364. [Google Scholar] [CrossRef]
- Lunkenbein, S.; Bellido, M.; Aharoni, A.; Salentijn, E.M.J.; Kaldenhoff, R.; Coiner, H.A.; Munoz-Blanco, J.; Schwab, W. Cinnamate metabolism in ripening fruit. Characterization of a UDP-glucose: Cinnamate glucosyltransferase from strawberry. Plant Physiol. 2006, 140, 1047–1058. [Google Scholar] [CrossRef]
- Gasperotti, M.; Masuero, D.; Guella, G.; Palmieri, L.; Martinatti, P.; Pojer, E.; Mattivi, F.; Vrhovsek, U. Evolution of ellagitannin content and profile during fruit ripening in Fragaria spp. J. Agric. Food Chem. 2013, 61, 8597–8607. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Guella, G.; Gasperotti, M.; Pojer, E.; Zancato, M.; Mattivi, F. Clarifying the identity of the main ellagitannin in the fruit of the strawberry, Fragaria vesca and Fragaria ananassa Duch. J. Agric. Food Chem. 2012, 60, 2507–2516. [Google Scholar] [CrossRef] [PubMed]
- Karlińska, E.; Masny, A.; Cieślak, M.; Macierzyński, J.; Pecio, Ł.; Stochmal, A.; Kosmala, M. Ellagitannins in roots, leaves, and fruits of strawberry (Fragaria × ananassa Duch.) vary with developmental stage and cultivar. Sci. Hortic. 2021, 275, 109665. [Google Scholar] [CrossRef]
- Pan, H.; Lundgren, L.N. Phenolic extractives from root bark of Picea abies. Phytochemistry 1995, 39, 1423–1428. [Google Scholar] [CrossRef]
- Yang, D.; Xie, H.; Jiang, Y.; Wei, X. Phenolics from strawberry cv. Falandi and their antioxidant and α-glucosidase inhibitory activities. Food Chem. 2016, 194, 857–863. [Google Scholar] [CrossRef]
- Li, W.; Shen, X.; Wang, J.; Sun, X.; Yuan, Q. Engineering microorganisms for the biosynthesis of dicarboxylic acids. Biotechnol. Adv. 2021, 48, 107710. [Google Scholar] [CrossRef]
- Yamagaki, T.; Watanabe, T. Hydrogen radical removal causes complex overlapping isotope patterns of aromatic carboxylic acids in negative-ion matrix-assisted laser desorption/ionization mass spectrometry. Mass Spectrom. 2012, 1, A0005. [Google Scholar] [CrossRef]
- Markham, K.R. Techniques of Flavonoids Identification; Academic Press: London, UK, 1982; ISBN 0-12-472680-1. [Google Scholar]
- D’Urso, G.; Pizza, C.; Piacente, S.; Montoro, P. Combination of LC-MS based metabolomics and antioxidant activity for evaluation of bioactive compounds in Fragaria vesca leaves from Italy. J. Pharm. Biomed. Anal. 2018, 150, 233–240. [Google Scholar] [CrossRef]
- Li, X.; Zheng, T.; Sang, S.; Lv, L. Quercetin inhibits advanced glycation end product formation by trapping methylglyoxal and glyoxal. J. Agric. Food Chem. 2014, 62, 12152–12158. [Google Scholar] [CrossRef]
- Brings, S.; Fleming, T.; Freichel, M.; Muckenthaler, M.; Herzig, S.; Nawroth, P. Dicarbonyls and advanced glycation end-products in the development of diabetic complications and targets for intervention. Int. J. Mol. Sci. 2017, 18, 984. [Google Scholar] [CrossRef]
- Van den Eynde, M.D.G.; Geleijnse, J.M.; Scheijen, J.L.J.M.; Hanssen, N.M.J.; Dower, J.I.; Afman, L.A.; Stehouwer, C.D.A.; Hollman, P.C.H.; Schalkwijk, C.G. Quercetin, but not epicatechin, decreases plasma concentrations of methylglyoxal in adults in a randomized, double-blind, placebo-controlled, crossover trial with pure flavonoids. J. Nutr. 2018, 148, 1911–1916. [Google Scholar] [CrossRef]
- Shatylo, V.; Antoniuk-Shcheglova, I.; Naskalova, S.; Bondarenko, O.; Havalko, A.; Krasnienkov, D.; Zabuga, O.; Kukharskyy, V.; Guryanov, V.; Vaiserman, A. Cardio-metabolic benefits of quercetin in elderly patients with metabolic syndrome. PharmaNutrition 2021, 15, 100250. [Google Scholar] [CrossRef]
- Yi, H.; Peng, H.; Wu, X.; Xu, X.; Kuang, T.; Zhang, J.; Du, L.; Fan, G. The therapeutic effects and mechanisms of quercetin on metabolic diseases: Pharmacological data and clinical evidence. Oxid. Med. Cell. Longev. 2021, 2021, 6678662. [Google Scholar] [CrossRef]
- Hai, Y.; Zhang, Y.; Liang, Y.; Ma, X.; Qi, X.; Xiao, J.; Xue, W.; Luo, Y.; Yue, T. Advance on the absorption, metabolism, and efficacy exertion of quercetin and its important derivatives. Food Front. 2020, 1, 420–434. [Google Scholar] [CrossRef]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of quercetin in humans with a focus on interindividual variation. Compr. Rev. Food Sci. Food Saf. 2018, 17, 714–731. [Google Scholar] [CrossRef]
- Van der Lugt, T.; Opperhuizen, A.; Bast, A.; Vrolijk, M.F. Dietary advanced glycation endproducts and the gastrointestinal tract. Nutrients 2020, 12, 2814. [Google Scholar] [CrossRef]
- Bhuiyan, M.N.I.; Mitsuhashi, S.; Sigetomi, K.; Ubukata, M. Quercetin inhibits advanced glycation end product formation via chelating metal ions, trapping methylglyoxal, and trapping reactive oxygen species. Biosci. Biotechnol. Biochem. 2017, 81, 882–890. [Google Scholar] [CrossRef]
- Van Wyk, B.-E.; Wink, M. Fragaria vesca wild strawberry. In Medicinal Plants of the World: An Illustrated Scientific Guide to Important Medicinal Plants and Their Uses; Briza Publications: Pretoria, South Africa, 2004; p. 146. ISBN 1-875093-44-3. [Google Scholar]
- Marunaka, Y. Actions of quercetin, a flavonoid, on ion transporters: Its physiological roles. Ann. N. Y. Acad. Sci. 2017, 1398, 142–151. [Google Scholar] [CrossRef]
- Vargas, F.; Romecín, P.; García-Guillén, A.I.; Wangesteen, R.; Vargas-Tendero, P.; Paredes, M.D.; Atucha, N.M.; García-Estañ, J. Flavonoids in kidney health and disease. Front. Physiol. 2018, 9, 394. [Google Scholar] [CrossRef]
- Wang, X.; Cao, Y.; Chen, S.; Lin, J.; Bian, J.; Huang, D. Anti-inflammation activity of flavones and their structure–activity relationship. J. Agric. Food Chem. 2021, 69, 7285–7302. [Google Scholar] [CrossRef]
- Dhanya, R. Quercetin for managing type 2 diabetes and its complications, an insight into multitarget therapy. Biomed. Pharmacother. 2022, 146, 112560. [Google Scholar] [CrossRef]
- Yong, Y.; Shin, S.Y.; Jung, Y.; Jung, H.; Ahn, S.; Chong, Y.; Lim, Y. Flavonoids activating adenosine monophosphate-activated protein kinase. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 13–19. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, Q.; Zhou, Y. Strawberry leaf extract treatment alleviates cognitive impairment by activating Nrf2/HO-1 signaling in rats with streptozotocin-induced diabetes. Front. Aging Neurosci. 2020, 12, 201. [Google Scholar] [CrossRef]
- Ibrahim, D.S.; Abd El-Maksoud, M.A.E. Effect of strawberry (Fragaria × ananassa) leaf extract on diabetic nephropathy in rats. Int. J. Exp. Pathol. 2015, 96, 87–93. [Google Scholar] [CrossRef]
- Kashchenko, N.; Chirikova, N.; Olennikov, D. Agrimoniin, an active ellagitannin from Comarum palustre herb with anti-α-glucosidase and antidiabetic potential in streptozotocin-induced diabetic rats. Molecules 2017, 22, 73. [Google Scholar] [CrossRef]
- Cardoso, O.; Donato, M.M.; Luxo, C.; Almeida, N.; Liberal, J.; Figueirinha, A.; Batista, M.T. Anti-Helicobacter pylori potential of Agrimonia eupatoria L. and Fragaria vesca. J. Funct. Foods 2018, 44, 299–303. [Google Scholar] [CrossRef]
- Juergenliemk, G.; Boje, K.; Huewel, S.; Lohmann, C.; Galla, H.-J.; Nahrstedt, A. In vitro studies indicate that miquelianin (quercetin 3-O-β-D-glucuronopyranoside) is able to reach the CNS from the small intestine. Planta Med. 2003, 69, 1013–1017. [Google Scholar]
- Albergamo, A.; Costa, R.; Bartolomeo, G.; Rando, R.; Vadalà, R.; Nava, V.; Gervasi, T.; Toscano, G.; Germanò, M.P.; D’Angelo, V.; et al. Grape water: Reclaim and valorization of a by-product from the industrial cryoconcentration of grape (Vitis vinifera) must. J. Sci. Food Agric. 2020, 100, 2971–2981. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Ibrahim, B.M.M.; Abdel-Latif, Y.; Hassan, A.H.; El Raey, M.A.; Hassan, E.M.; El-Gengaihi, S.E. Pharmacological and metabolomic profiles of Musa acuminata wastes as a new potential source of anti-ulcerative colitis agents. Sci. Rep. 2022, 12, 10595. [Google Scholar] [CrossRef]
- Petrović, M.; Jovanović, M.; Lević, S.; Nedović, V.; Mitić-Ćulafić, D.; Živković Semren, T.; Veljović, S. Valorization potential of Plantago major L. solid waste remaining after industrial tincture production: Insight into the chemical composition and bioactive properties. Waste Biomass Valorization 2022, 13, 1639–1651. [Google Scholar] [CrossRef]
- Slavov, A.; Yantcheva, N.; Vasileva, I. Chamomile wastes (Matricaria chamomilla): New source of polysaccharides. Waste Biomass Valorization 2019, 10, 2583–2594. [Google Scholar] [CrossRef]
- Mitharwal, S.; Kumar, A.; Chauhan, K.; Taneja, N.K. Nutritional, phytochemical composition and potential health benefits of taro (Colocasia esculenta L.) leaves: A review. Food Chem. 2022, 383, 132406. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Pratibha, N.; Sami, R.; Khojah, E.; Aljahani, A.H.; Al-Mushhin, A.A.M. Effects of drying methods and solvent extraction on quantification of major bioactive compounds in pomegranate peel waste using HPLC. Sci. Rep. 2022, 12, 8000. [Google Scholar] [CrossRef] [PubMed]
- Palos-Hernandez, A.; Gutierrez Fernandez, M.Y.; Escuadra Burrieza, J.; Perez-Iglesias, J.L.; Gonzalez-Paramas, A.M. Obtaining green extracts rich in phenolic compounds from underexploited food by-products using natural deep eutectic solvents. Opportunities and challenges. Sustain. Chem. Pharm. 2022, 29, 100773. [Google Scholar] [CrossRef]
- Tao, W.; Jin, J.; Zheng, Y.; Li, S. Current advances of resource utilization of herbal extraction residues in China. Waste Biomass Valorization 2021, 12, 5853–5868. [Google Scholar] [CrossRef]
- Virbala, S.; Shreekar, P. Weed as underutilized bio-resource and management tool: A comprehensive review. Waste Biomass Valorization 2019, 10, 1795–1810. [Google Scholar]
- Fecka, I.; Włodarczyk, M.; Starzec, A. Isolation and structure elucidation of cistusin: A new ellagitannin from Cistus × incanus L. leaves. Ind. Crops Prod. 2020, 158, 112971. [Google Scholar] [CrossRef]
- Tanaka, T.; Nonaka, G.-I.; Nishioka, I. Tannins and related compounds. XLII. Isolation and characterization of four new hydrolyzable tannins, terflavins A and B, tergallagin and tercatain from the leaves of Terminalia catappa L. Chem. Pharm. Bull. (Tokyo) 1986, 34, 1039–1049. [Google Scholar] [CrossRef]
- Litvinienko, V.I.; Makarov, V.A. The alkaline hydrolysis of flavonoid glycosides. Chem. Nat. Compd. 1969, 5, 305–306. [Google Scholar] [CrossRef]
- Liu, W.; Ma, H.; Frost, L.; Yuan, T.; Dain, J.A.; Seeram, N.P. Pomegranate phenolics inhibit formation of advanced glycation endproducts by scavenging reactive carbonyl species. Food Funct. 2014, 5, 2996–3004. [Google Scholar] [CrossRef]
- Sang, S.; Shao, X.; Bai, N.; Lo, C.-Y.; Yang, C.S.; Ho, C.-T. Tea polyphenol (−)-epigallocatechin-3-gallate: A new trapping agent of reactive dicarbonyl species. Chem. Res. Toxicol. 2007, 20, 1862–1870. [Google Scholar] [CrossRef]
- Friedrich, W.; Eberhardt, A.; Galensa, R. Investigation of proanthocyanidins by HPLC with electrospray ionization mass spectrometry. Eur. Food Res. Technol. 2000, 211, 56–64. [Google Scholar] [CrossRef]
- Hamed, A.I.; Al-Ayed, A.S.; Moldoch, J.; Piacente, S.; Oleszek, W.; Stochmal, A. Profiles analysis of proanthocyanidins in the argun nut (Medemia argun—An ancient Egyptian palm) by LC-ESI-MS/MS. J. Mass Spectrom. 2014, 49, 306–315. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tR [min] | UV λmax [nm] | MS1[M−H]− [m/z] Measured | MS1[M−H]− [m/z] Calculated | MS2[M−H]− [m/z] Measured | Identification | Compound, Reference |
|---|---|---|---|---|---|---|
| Carboxylic acids | ||||||
| 1.76 | 315 | 182.9934, 366.9940 [2M−H]− | 182.9935 | 139 [M−44/CO2−H]−, 111 [M−44/CO2−28/CO−H]− | 2-pyrone-4,6-dicarboxylic acid, PDC | (1), isol, [25] |
| 1.32 | - | 191.0198 | 191.0197 | 111 | citric acid (isomers) | std |
| 1.45 | ||||||
| 1.15 | - | 191.0563 | 191.0561 | 173 [M−18/H2O−H]−, 155 [M−36/2H2O−H]−, 111 [M−44/CO2−36/2H2O−H]− | quinic acid | (2), std |
| Hydrolyzable tannins | ||||||
| 7.24 | 275 | 331.0669 | 331.0671 | 169 [GA−H]−, 125 [GA−44/CO2−H]− | galloyl-glucose, e.g., 1-O-galloyl-α-glucose | [27,28] |
| 1.21 | 273 | 343.0669, 687.1407 [2M−H]− | 343.0671 | 191 [QA−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | O-galloylquinic acid (isomer 1) | [27,28] |
| 1.52 | 3-O-galloylquinic acid (isomer 2) | (3), isol, [14] | ||||
| 3.02 | 191 [QA−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | 5-O-galloylquinic acid, theogallin (isomer 3) | (5), isol, [14] | |||
| 3.74 | 191 [QA−H]−, 173 [QA−18/H2O−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | O-galloylquinic acid (isomer 4) | [14] | |||
| 11.75 | 275 | 483.0770 | 483.0780 | 169 [GA−H]−, 125 [GA−44/CO2−H]− | digalloyl-glucose | [27,28] |
| 11.19 | 273 | 495.0778 | 495.0780 | 343 [M−152/gall−H]−, 191 [QA−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | 3,5-O-digalloylquinic acid (or isomer) | [14] |
| 9.03 | 242, 275 | 633.0724 | 633.0733 | 481 [M−152/gall−H]−, 463 [M−152/gall−18/H2O−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | galloyl-HHDP-glucose (isomers), e.g., sanguiin H-4 | [27,28] |
| 10.79 | ||||||
| 13.54 | 275 | 635.0894 | 635.0890 | 465 [M−152/gall−18/H2O−H]−, 313 [M−304/2gall−18/H2O−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | trigalloylglucose | [27,28] |
| 2.88 | 243 | 783.0676 | 783.0686 | 481 [M−302/HHDP−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]− | pedunculagin (isomers), e.g., α/β anomers | (7a) and (7b), isol, [26,29,30] |
| 5.36 | ||||||
| 8.68 | ||||||
| 10.20 | 273 | 785.0826 | 785.0843 | 633 [M−152/gall−H]−, 481 [M−304/2gall−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | digalloyl-HHDP glucose | [28] |
| 11.81 | 345 | 933.0656 466.0270 [M−2H]−2 | 933.0640, 466.0283 [M−2H]− | 631[M−302/HHDP−H]−, 481 [M−452/VTL−H]−, 451 [VTL−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, [GA−H]−, 125 [GA−44/CO2−H]− | ellagitannin-934 (isomers), e.g., α/β anomers of praecoxin D | [30,31] |
| 12.98 | ||||||
| 13.17 | 275 | 935.0808, 467.0354 [M−2H]−2 | 935.0796, 467.0362 [M−2H]− | 783 [M−152/gall−H]−, 633 [M−302/HHDP−H]−, 613 [M−304/2gall−18/H2O−H]−, 481 [M−302/HHDP−152/gall−H]−, 463 [M−302/HHDP−152/gall−18/H2O−H]−, 313 [M−604/2HHDP−18/H2O−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | potentillin | [28,30] |
| 14.47 | 243 | 1235.0744, 617.0299 [M−2H]−2 | 1235.0702, 617.0315 [M−2H]−2 | 933 [M−302/HHDP or DHDG−H]−, 935 [M−300−H]− (potentillin), 613 [M−302/HHDP−302/DHDG−18/H2O−H]−, 783 [M−452/VTL−H]−, 481 [M−452/VTL−302/HHDP or DHDG−H]−, 469 [val−H]−, 463 [M−302/HHDP or DHDG−470/val−H]−, 451 [VTL−H]−, 319 [LDHDG−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, [GA−H]−, 125 [GA−44/CO2−H]− | ellagitannin-1236, e.g., davuriciin D2 / fragariin A lactone without a pedunculagin fragment | [28,30,32] |
| 12.31 | 245, 276 | 783.0672 [M−2H]−2 | 1567.1446, 783.0686 [M−2H]−2 | 1265 [M−302/HHDP−H]− (laevigatin E), 1103 [M−302/HHDP−162/glc−H]− (agrimonic acid A/B), 1059 [M−302/HHDP−162/glc−44/CO2−H]−, 935 [M−632−H]− (potentillin), 783 [M−784/pedunculagin−H]− (pedunculagin or laevigatin A lctone), 633 [M−934−H]−, 613 [M−936−18/H2O−H]−, 481 [M−1086 /agrimonic acid lactone−H]−, 463 [M−1086/agrimonic acid lactone−18/H2O−H]−, 319 [LDHDG−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, 125 [GA−44/CO2−H]− | laevigatin B/C/F (isomers) | [33,34,35] |
| 13.17 | ||||||
| 13.93 | ||||||
| 14.33 | 245 | 934.0722 [M−2H]−2 | 1869.1508, 934.0718 [M−2H]−2 | 1567 [M−302/HHDP−H]− (laevigatin B/C/F), 1085 [M−784/ pedunculagin−H]−, 935 [M−934−H]− (potentillin), 897 [M−936/ potentillin−36/2H2O−H]−, 783 [M−1086/agrimonic acid A/B lactone−H]− (pedunculagin) or [M−784/ pedunculagin−302/HHDP−H]− (laevigatin A lactone), 745 [M−936/potentillin−152/gall− 36/2H2O)−H]−, 633 [M−1236−H]−, 613 [M−1086/agrimonic acid lactone A/B−152/gall−18/H2O−H]−, 481 [M−1086/agrimonic acid A/B lactone−302/HHDP−H]−, 463 [M−1086(agrimonic acid A/B lactone)−302(HHDP)−18(H2O)−H]−, 319 [LDHDG−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, 125 [GA−44/CO2−H]− | agrimoniin | (10), isol, [11,29,33] |
| 16.99 | 243 | 1009.0686 [M−2H]−2 | 2019.1461, 1009.0694 [M−2H]−2 | 1235 [M−784/pedunculagin−H]−, 933 [M−784/pedunculagin-302/DHDG−H]− (praecoxin D), 897 [M−1104/agrimonic acid B−18/H2O−H]−, 783 [M−1236−H]− (pedunculagin or laevigatin A lactone), 769 [M−1252−H]−, 633 [M−934−452/VTL−H]− or [M−1084−302/HHDP−H]−, 613 [M−1104/agrimonic acid B-302/ HHDP−H]−, 481 [M−1086/agrimonic acid lactone−452/VTL−H]−, 451 [VTL−H]−, 319 [LDHDG−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, 169 [GA−H]−, 125 [GA−H]− | ellagitannin-2020, e.g., davuriciin D2/fragariin A lactone | [27,28,36] |
| 14.23 | 244 | 1018.0749 [M−2H]−2 | 2037.1567, 1018.0747 [M−2H]−2 | 1567 [M−470/val−H]− (laevigatin B/C/F), 933 [M−1104/agrimonic acid B−H]−, 783 [M−1236−18/H2O−H]− (pedunculagin or laevigatin A lactone), 745 [M−1104/ agrimonic acid B−152/gall−36/2H2O−H]−, 633 [M−934−452/VTL−18/H2O−H]− or [M−1084−302/HHDP−18/H2O−H]−, 613 [M−1236−152/gall−36/2H2O−H]−, 481 [M−1086−470/val−H]−, 463 [M−1086−470/val−18/H2O−H]−, 451 [VTL−H]−, 319 [LDHDG−H]−, 301 [EA−H]−, 275 [LHHDP−44/CO2−H]−, 169 [GA−H]−, 125 [GA−44/CO2−H]− | davuriciin D2/fragariin A | [27,28,32,36,37] |
| Proanthocyanidins (condensed tannins) and flavan−3−ols | ||||||
| 10.86 | 242, 278 | 289.0719 | 289.0712 | 245 [C13H10O5−H]−, 205 [C11H10O4−H]−, 203 [C11H8O4−H]−, 151 [C8H8O3−H]−,137 [C7H6O3−H]−, 125 [C6H6O3−H]−, 123 [C7H8O2−H]−, 121 [C7H6O2−H]−, 109 [C6H6O2−H]− | catechin | (6), isol, std, [26] |
| 15.55 | 242, 278 | 451.1237 | 451.1246 | 289 [M−162/hex−H]− | (epi)catechin-O-hexoside | - |
| 9.78 | 242, 278 | 577.1346 | 577.1352 | 451 [M−126/C6H6O3−H]−, 425 [M−152/C8H8O3−H]−, 407 [M−152/ C8H8O3−18/H2O−H]−, 289 [M−288/C15H12O6−H]−, 287 [M−290/ C15H14O6−H]−, 245 [C13H10O5−H]−, 137 [C7H6O3−H]−, 125 [C6H6O3−H]− | catechin dimer, e.g., procyanidin B3 | std, [26,33] |
| 10.13 | 425 [M−152/C8H8O3−H]−, 407 [M−152/C8H8O3−18/H2O−H]−, 289 [M−288/C15H12O6−H]−, 287 [M−290/C15H14O6−H]−, 137 [C7H6O3−H]−, 125 [C6H6O3−H]− | (epi)catechin dimer (isomers) | [27,28,30,36] | |||
| 10.23 | ||||||
| 11.77 | ||||||
| 13.22 | ||||||
| 10.13 | 242, 278 | 865.1981 | 865.1985 | 739 [M−126/C6H6O3−H]−, 713 [M−152/C8H8O3−H]−, 695 [M−152/ C8H8O3−18/H2O−H]−, 577 [M−288/C15H12O6−H]−, 575 [M−290/ C15H14O6−H]−, 451 [M−288/C15H12O6−126/ C6H6O3−H]−, 425 [M−288/C15H12O6−152/C8H8O3−H]−, 407 [M−288/C15H12O6−152/C8H8O3−18/H2O−H]−, 289 [M−576/C30H24O12−H]−, 287 [M−578/C30H26O12−H]−, 245 [C13H10O5−H]−, 137 [C7H6O3−H]−, 125 [C6H6O3−H]− | catechin trimer, e.g., procyanidin C3 | std, [33] |
| 10.66 | (epi)catechin trimer (isomers) | [27,28,30,33,36] | ||||
| 11.49 | ||||||
| 10.92 | 242, 278 | 1153.2610 | 1153.2619 | 1027 [M−126/C6H6O3−H]− 1001 [M−152/C8H8O3−H]−, 983 [M−152/ C8H8O3−18/H2O−H]−, 865 [M−288/C15H12O6−H]−, 863 [M−290/ C15H14O6−H]−, 695 [M−288/C15H12O6−152/ C8H8O3−18/H2O−H]−, 577 [M−576/C30H24O12−H]−, 575 [M−578/C30H26O12−H]−, 451 [M−576/C30H24O12−126/C6H6O3−H]−, 425 [M−576/C30H24O12− 152/C8H8O3−H]−, 407 [M−576/ C30H24O12−152/C8H8O3−18/H2O−H]−, 289 [M−864/C45H36O18−H]−, 287 [M−866/C45H38O18−H]−, 137 [C7H6O3−H]−, 125 [C6H6O3−H]− | (epi)catechin tetramer | |
| Flavonoids | ||||||
| 17.08 | 290 | 435.0930 | 435.0933 | 303 [M−132/pent−H]−, 285 [M−162/glc−18/H2O−H]−, 179, 151 | taxifolin-O-pentoside (isomers), e.g., taxifolin-3-O-α-arabinoside | [26] |
| 21.03 | ||||||
| 19.23 | 265, 345 | 447.0929 | 447.0933 | 284/285 [M−162/glc−H]−, 229 | kaempferol-3-O-β-glucoside (astragalin) | std, [21] |
| 19.19 | 265, 345 | 461.0724 | 461.0726 | 285 [M−176/glcA−H]−, 255, 229, 151 | kaempferol-3-O-β-glucuronoside | std, [28,36] |
| 17.91 | 255, 353 | 463.0878 | 463.0882 | 301 [M−162/glc−H]−, 179, 151 | quercetin-3-O-β-glucoside (isoquercitrin) | std |
| 10.61 | 226, 289 | 465.1035 | 465.1039 | 303 [M−162/glc−H]−, 285 [M−162/glc−18/H2O−H]−, 151 | taxifolin-O-hexoside (isomer 1) | [21] |
| 14.23 | (2R,3R)-taxifolin 3-O-β-glucoside (isomer 2) | (8), isol, [21] | ||||
| 15.18 | taxifolin-O-hexoside (isomer 3) | [21] | ||||
| 15.77 | taxifolin-O-hexoside (isomer 4) | |||||
| 17.74 | 255, 353 | 477.0672 | 477.0675 | 301 [M−176/glcA−H]−, 179, 151 | quercetin-3-O-β-glucuronoside (miquelianin) | (12), isol, [21,28,36] |
| 19.18 | 239, 255, 353 | 491.0806 | 491.0831 | 315 [M−176/glcA−H]−, 300 [M−176−15/Me•−H]•−, 137 | isorhamnetin-3-O-β-glucuronoside | (14), isol, [28,38] |
| 17.94 | 265, 347 | 593.1144 | 593.1148 | 285 [M−308−H]−, 229, 151 | kaempferol-3-O-[β-xylosyl(1‴→2″)]-β-glucuronoside | (13), isol, [27,28] |
| 22.07 | 264, 312, 347 | 593.1299 | 593.1300 | 447 [M−146/p-coumaroyl−H]−, 285 [M−146/p-coumaroyl−162/glc−H]−, 145 [p-coumaroyl−H]−, 119 [pCouA−44/CO2−H]− | tiliroside (cis/trans isomers) | (15a) and (15b), isol, [39,40,41,42] |
| 22.25 | ||||||
| 16.59 | 256, 353 | 609.1108 | 609.1097 | 301 [M−308−H]−, 179, 151 | quercetin-3-O-[β-xylosyl(1‴→2″)]-β-glucuronoside, (flagarin) | (11), isol, [21,27,28] |
| 13.01 | 256, 352 | 639.1200 | 639.1203 | 463 [M−176/glcA−H]−, 301 [M−176−162/glc−H]−, 179, 151 | quercetin-3-O-β-glucuronoside-7-O-β-glucoside | (9), isol, [28] |
| 15.77 | 255, 351 | 301 [M−338−H]−, 179, 151 | quercetin-3-O-(β-glucosyl)-β-glucuronoside | [28] | ||
| 9.34 | 227 | 285.0614 | 285.0616 | 152/153 [M−132/xyl−H]−, 108/109 [PA−44/CO2−H]− | 1-O-protocatechuoyl-β-xylose | (4a), isol |
| Phenolic acids’ derivatives | ||||||
| 18.18 | 253, 367 | 300.9987 | 300.9990 | 301 [EA−H]−, 275 [LHHDP−44/CO2−H]− | ellagic acid | (16), isol, std |
| 6.14 | 282 | 315.0718 | 315.0722 | 153 [M−162/glc−H]−, 109 [PA−44/CO2−H]− | protocatechuic acid 3-O-β-glucoside | (4b), isol |
| 11.35 | 320 | 325.0927 | 325.0929 | 163 [M−162/hex−H]−, 145 [p-coumaroyl−H]−, 119 [pCuA−44/CO2−H]− | pCuA hexoside or ester (isomers), e.g., p-coumaric acid 4-O-β-glucoside, 1-O-p-coumaroyl-β-glucose | [21,28,36,43] |
| 11.81 | ||||||
| 12.21 | ||||||
| 13.13 | ||||||
| 14.54 | ||||||
| 19.20 | ||||||
| 8.14 | 322 | 341.0878 | 341.0878 | 179 [M−162/hex−H]−, 161 [caffeoyl−H]−, 135 [CA−44/CO2−H]− | CA hexoside or ester (isomers), e.g., 1-O-caffeoyl-β-glucose | [43] |
| 9.03 | ||||||
| 10.64 | ||||||
| 11.04 | ||||||
| 11.42 | ||||||
| 12.62 | ||||||
| 7.57 | 322 | 355.0665 | 355.0671 | 179 [M−176/glcA−H]−, 161 [caffeoyl−H]−, 135 [CA−44/CO2−H]− | CA glucuronoside or ester (isomers) | |
| 8.94 | ||||||
| 9.41 | ||||||
| 10.30 | ||||||
| 11.09 | ||||||
| 12.65 | ||||||
| 17.48 | 253, 361 | 433.0408 | 433.0412 | 301 [EA−132/pent−H]−, 275 [LHHDP−44/CO2−H]− | ellagic acid O-pentoside | [28,36] |
| 19.50 | 246, 312 | 445.1507 | 445.1504 | 353, 236, 205, 163 [pCuA−H]−, 145 [p-coumaroyl−H]−, 119 [pCuA−44/CO2−H]− | eutigoside A (isomers) | [28] |
| 19.78 | ||||||
| 18.01 | 245, 377 | 447.0570 | 447.0569 | 301 [M−146/dhex−H]−, 275 [LHHDP−44/CO2−H]− | ellagic acid O-deoxyhexoside | [28,36,44] |
| 18.12 | 246, 285, 323 | 461.1448 | 461.1453 | 179 [M−162/hex−H]−, 161 [M−162/hex−138/hydroxyphenylethanol−H]−, 135 [CA−44/CO2−H]− | O-hydroxyphenylethyl-O-caffeoyl-glucoside | [42] |
| Compound | Abbreviation | FaSS1 | FaSS2 | FaSS3 | |||
|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | ||
| 2-pyrone-4,6-dicarboxylic acid (1) | PDC | 13.73 | 0.31 | 10.37 | 0.28 | 17.45 | 0.45 |
| 5-O-galloylquinic acid (5) | 5gQA | 5.21 | 0.27 | 4.10 | 0.16 | 6.43 | 0.30 |
| agrimoniin (10) | A | 44.50 | 1.80 | 29.87 | 0.87 | 80.82 | 3.29 |
| davuriciin D2/fragariin A 1 | D2 1 | 6.43 | 0.19 | 2.06 | 0.13 | 7.64 | 0.19 |
| Sum of ellagitannins | ETs | 50.97 | 1.51 | 31.93 | 0.82 | 88.42 | 2.42 |
| quercetin-3-O-[β-D-xylosyl-(1‴→2″)]-β-D-glucuronoside (11) | Q3grx | 7.27 | 0.28 | 6.94 | 0.10 | 7.21 | 0.18 |
| quercetin-3-O-β-D-glucuronoside-7-O-β-D-glucoside (9) | Q3gr7g | 0.73 | 0.03 | 1.07 | 0.03 | 0.83 | 0.06 |
| quercetin-3-O-β-D-glucuronoside (12) | Q3gr | 3.39 | 0.09 | 1.65 | 0.02 | 1.46 | 0.12 |
| kaempferol-3-O-[β-D-xylosyl-(1‴→2″)]-β-D-glucuronoside (13) | K3grx | 5.09 | 0.06 | 4.86 | 0.22 | 5.86 | 0.12 |
| kaempferol-3-O-β-D-glucuronoside | K3gr | 0.57 | 0.01 | 0.51 | 0.01 | 0.42 | 0.04 |
| Sum of flavonols | Fs | 17.06 | 0.09 | 15.02 | 0.08 | 15.77 | 0.10 |
| Compound | tR [min] | [M−H]− [m/z], Measured | Peak/Adducts |
|---|---|---|---|
| quercetin (Q) | 10.01 | 445.0779 | Di-MGO-Q |
| 10.93; 11.06 | 373.0567 | Mono-MGO-Q (two isomers) | |
| 11.83 | 301.0355 | Q | |
| quercetin-3-O-β-D-glucoside isoquercitrin (Q3g) | 8.78; 8.87; 9.00; 9.12 | 535.1106 | Mono-MGO-Q3g (four isomers) |
| 9.95 | 463.0890 | Q3g | |
| quercetin-4′-O-β-D-glucoside spiraeoside (Q4′g) | 8.98 | 607.1279 | Di-MGO-Q4′g |
| 9.61 | 535.1068 | Mono-MGO-Q4′g | |
| 10.67 | 463.0856 | Q4′g | |
| quercetin-3-O-β-D-galactoside hyperoside (Q3ga) | 8.19; 8.28; 8.37 | 607.1303 | Di-MGO-Q3ga (three isomers) |
| 8.70; 8.79; 8.99; 9.09 | 535.1099 | Mono-MGO-Q3ga (four isomers) | |
| 9.88 | 463.0894 | Q3ga | |
| quercetin-3-O-β-D-glucuronoside miquelianin (12; Q3gr) | 8.21; 8.48; 8.68 | 621.1082 | Di-MGO-Q3gr (three isomers) |
| 8.83; 9.04; 9.13; 9.31 | 549.0872 | Mono-MGO-Q3gr (four isomers) | |
| 10.20 | 477.0672 | Q3gr | |
| quercetin-3-O-[β-D-xylosyl(1‴→2″)]-β-D-glucuronoside flagarin (11; Q3grx) | 8.09; 8.46; 8.75 | 753.1536 | Mono-MGO-Q3grx (three isomers) |
| 9.45 | 609.1108 | Q3grx |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fecka, I.; Bednarska, K.; Włodarczyk, M. Fragaria × ananassa cv. Senga Sengana Leaf: An Agricultural Waste with Antiglycation Potential and High Content of Ellagitannins, Flavonols, and 2-Pyrone-4,6-dicarboxylic Acid. Molecules 2022, 27, 5293. https://doi.org/10.3390/molecules27165293
Fecka I, Bednarska K, Włodarczyk M. Fragaria × ananassa cv. Senga Sengana Leaf: An Agricultural Waste with Antiglycation Potential and High Content of Ellagitannins, Flavonols, and 2-Pyrone-4,6-dicarboxylic Acid. Molecules. 2022; 27(16):5293. https://doi.org/10.3390/molecules27165293
Chicago/Turabian StyleFecka, Izabela, Katarzyna Bednarska, and Maciej Włodarczyk. 2022. "Fragaria × ananassa cv. Senga Sengana Leaf: An Agricultural Waste with Antiglycation Potential and High Content of Ellagitannins, Flavonols, and 2-Pyrone-4,6-dicarboxylic Acid" Molecules 27, no. 16: 5293. https://doi.org/10.3390/molecules27165293