
Effect of Hypertrophic Scar Fibroblast-Derived Exosomes on Keratinocytes of Normal Human Skin
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Expression of Exosome Markers
2.2. Effects of NF- and HTSF-Exosomes on the Proliferation of Normal Human Keratinocytes (NHKs)
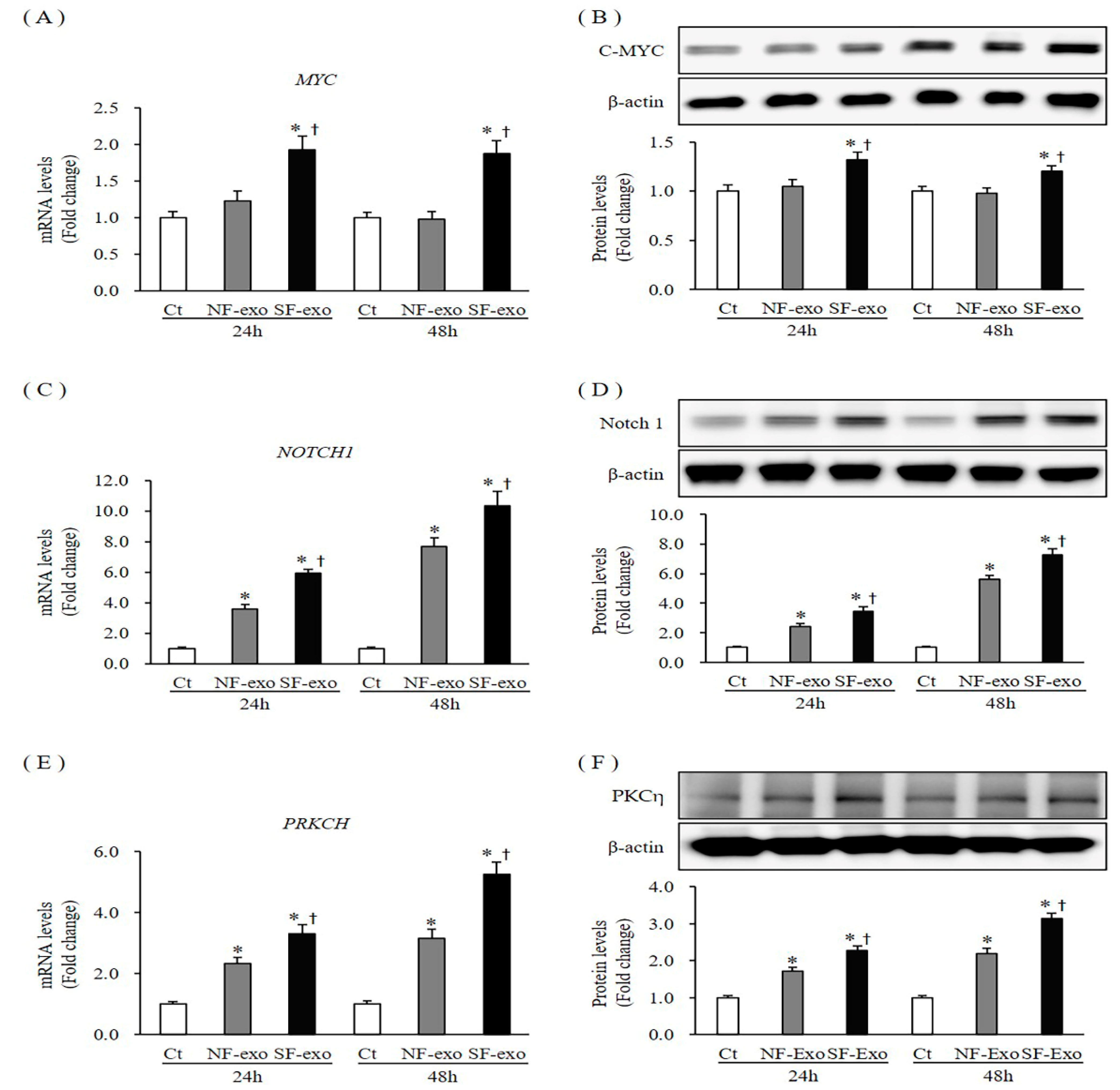
2.3. Effects of NF- and HTSF- Exosomes on Expression of Activation Markers in NHKs
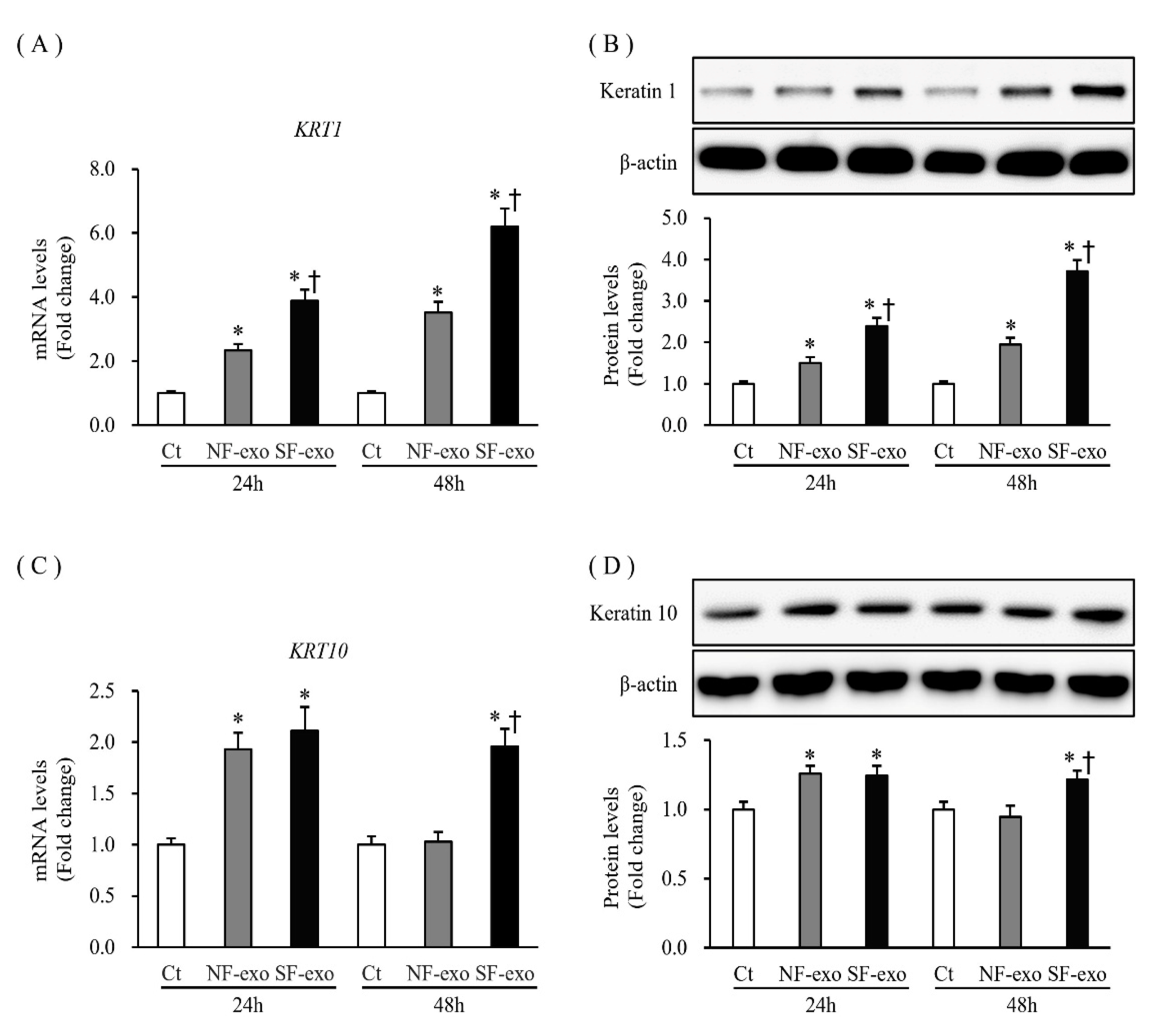
2.4. Effects of NF- and HTSF-Exosomes on the Expression of Differentiation Markers in NHKs
2.5. Effects of NF- and HTSF-Exosomes on the Expression of Apoptosis-Related Factors Related to Proliferation and Differentiation in NHKs
2.6. Effects of NF- and HTSF-Exosomes on the Expression of Cyclin-Dependent Kinase Inhibitors and Their Regulators in NHKs
2.7. Effects of NF- and HTSF-Exosomes on the Epithelial–Mesenchymal Transition (EMT) in NHKs
3. Discussion
4. Materials and Methods
4.1. Primary Cell Culture
4.2. Exosome Extraction and Treatment
4.3. Cell Proliferation Assay
4.4. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
4.5. Western Blot Analysis
4.6. Cell Migration Assay
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gauglitz, G.G.; Korting, H.C.; Pavicic, T.; Ruzicka, T.; Jeschke, M.G. Hypertrophic scarring and keloids: Pathomechanisms and current and emerging treatment strategies. Mol. Med. 2011, 17, 113–125. [Google Scholar] [CrossRef]
- Zhu, Z.; Ding, J.; Tredget, E.E. The molecular basis of hypertrophic scars. Burns Trauma 2016, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Tredget, E.E.; Shankowsky, H.A.; Pannu, R.; Nedelec, B.; Iwashina, T.; Ghahary, A.; Taerum, T.V.; Scott, P.G. Transforming growth factor-beta in thermally injured patients with hypertrophic scars: Effects of interferon alpha-2b. Plast. Reconstr. Surg. 1998, 102, 1317–1328, discussion 1329–1330. [Google Scholar] [CrossRef]
- Armour, A.; Scott, P.G.; Tredget, E.E. Cellular and molecular pathology of HTS: Basis for treatment. Wound Repair Regen. 2007, 15 (Suppl. 1), S6–S17. [Google Scholar] [CrossRef]
- Cui, H.S.; Hong, A.R.; Kim, J.B.; Yu, J.H.; Cho, Y.S.; Joo, S.Y.; Seo, C.H. Extracorporeal shock wave therapy alters the expression of fibrosis-related molecules in fibroblast derived from human hypertrophic scar. Int. J. Mol. Sci. 2018, 19, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penn, J.W.; Grobbelaar, A.O.; Rolfe, K.J. The role of the TGF-β family in wound healing, burns and scarring: A review. Int. J. Burns Trauma 2012, 2, 18–28. [Google Scholar]
- Moulin, V.; Larochelle, S.; Langlois, C.; Thibault, I.; Lopez-Vallé, C.A.; Roy, M. Normal skin wound and hypertrophic scar myofibroblasts have differential responses to apoptotic inductors. J. Cell. Physiol. 2004, 198, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Wassermann, R.J.; Polo, M.; Smith, P.; Wang, X.; Ko, F.; Robson, M.C. Differential production of apoptosis-modulating proteins in patients with hypertrophic burn scar. J. Surg. Res. 1998, 75, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in wound healing: A comprehensive review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [Green Version]
- Bader, R.A.; Kao, W.J. Modulation of the keratinocyte-fibroblast paracrine relationship with gelatin-based semi-interpenetrating networks containing bioactive factors for wound repair. J. Biomater. Sci. Polym. Ed. 2009, 20, 1005–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karvinen, S.; Pasonen-Seppänen, S.; Hyttinen, J.M.; Pienimäki, J.P.; Törrönen, K.; Jokela, T.A.; Tammi, M.I.; Tammi, R. Keratinocyte growth factor stimulates migration and hyaluronan synthesis in the epidermis by activation of keratinocyte hyaluronan synthases 2 and 3. J. Biol. Chem. 2003, 278, 49495–49504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallucci, R.M.; Sloan, D.K.; Heck, J.M.; Murray, A.R.; O’Dell, S.J. Interleukin 6 indirectly induces keratinocyte migration. J. Investig. Dermatol. 2004, 122, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, B.Z.; Stevenson, A.W.; Prêle, C.M.; Fear, M.W.; Wood, F.M. The role of IL-6 in skin fibrosis and cutaneous wound healing. Biomedicines 2020, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Campaner, A.B.; Ferreira, L.M.; Gragnani, A.; Bruder, J.M.; Cusick, J.L.; Morgan, J.R. Upregulation of TGF-beta1 expression may be necessary but is not sufficient for excessive scarring. J. Investig. Dermatol. 2006, 126, 1168–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Zavadil, J.; Bitzer, M.; Liang, D.; Yang, Y.C.; Massimi, A.; Kneitz, S.; Piek, E.; Bottinger, E.P. Genetic programs of epithelial cell plasticity directed by transforming growth factor-beta. Proc. Natl. Acad. Sci. USA 2001, 98, 6686–6691. [Google Scholar] [CrossRef] [Green Version]
- Liarte, S.; Bernabé-García, Á.; Nicolás, F.J. Human skin keratinocytes on sustained TGF-β stimulation reveal partial EMT features and weaken growth arrest responses. Cells 2020, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Amendt, C.; Mann, A.; Schirmacher, P.; Blessing, M. Resistance of keratinocytes to TGFbeta-mediated growth restriction and apoptosis induction accelerates re-epithelialization in skin wounds. J. Cell Sci. 2002, 115, 2189–2198. [Google Scholar] [CrossRef]
- Ding, J.; Hori, K.; Zhang, R.; Marcoux, Y.; Honardoust, D.; Shankowsky, H.A.; Tredget, E.E. Stromal cell-derived factor 1 (SDF-1) and its receptor CXCR4 in the formation of postburn hypertrophic scar (HTS). Wound Repair Regen. 2011, 19, 568–578. [Google Scholar] [CrossRef]
- Quan, C.; Cho, M.K.; Shao, Y.; Mianecki, L.E.; Liao, E.; Perry, D.; Quan, T. Dermal fibroblast expression of stromal cell-derived factor-1 (SDF-1) promotes epidermal keratinocyte proliferation in normal and diseased skin. Protein Cell 2015, 6, 890–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Charrier, A.; Chen, R.; Chen, L.; Kemper, S.; Hattori, T.; Takigawa, M.; Brigstock, D.R. Exosomes mediate intercellular transfer of pro-fibrogenic connective tissue growth factor (CCN2) between hepatic stellate cells, the principal fibrotic cells in the liver. Surgery 2014, 156, 548–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, F.T.; Melo, S.A.; Özdemir, B.C.; Kato, N.; Revuelta, I.; Miller, C.A.; Gattone, V.H.; LeBleu, V.S.; Kalluri, R. TGF-β1-containing exosomes from injured epithelial cells activate fibroblasts to initiate tissue regenerative responses and fibrosis. J. Am. Soc. Nephrol. 2013, 24, 385–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjan, P.; Kumari, R.; Goswami, S.K.; Li, J.; Pal, H.; Suleiman, Z.; Cheng, Z.; Krishnamurthy, P.; Kishore, R.; Verma, S.K. Myofibroblast-derived exosome induce cardiac endothelial cell dysfunction. Front. Cardiovasc. Med. 2021, 8, 676267. [Google Scholar] [CrossRef]
- Zanotti, S.; Gibertini, S.; Blasevich, F.; Bragato, C.; Ruggieri, A.; Saredi, S.; Fabbri, M.; Bernasconi, P.; Maggi, L.; Mantegazza, R.; et al. Exosomes and exosomal miRNAs from muscle-derived fibroblasts promote skeletal muscle fibrosis. Matrix Biol. 2018, 74, 77–100. [Google Scholar] [CrossRef]
- Cui, H.S.; Kim, D.H.; Joo, S.Y.; Cho, Y.S.; Kim, J.B.; Seo, C.H. Exosomes derived from human hypertrophic scar fibroblasts induces smad and TAK1 signaling in normal dermal fibroblasts. Arch. Biochem. Biophys. 2022, 722, 109215. [Google Scholar] [CrossRef] [PubMed]
- Usui, M.L.; Mansbridge, J.N.; Carter, W.G.; Fujita, M.; Olerud, J.E. Keratinocyte migration, proliferation, and differentiation in chronic ulcers from patients with diabetes and normal wounds. J. Histochem. Cytochem. 2008, 56, 687–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedberg, I.M.; Tomic-Canic, M.; Komine, M.; Blumenberg, M. Keratins and the keratinocyte activation cycle. J. Investig. Dermatol. 2001, 116, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Missero, C.; Di Cunto, F.; Kiyokawa, H.; Koff, A.; Dotto, G.P. The absence of p21Cip1/WAF1 alters keratinocyte growth and differentiation and promotes ras-tumor progression. Genes Dev. 1996, 10, 3065–3075. [Google Scholar] [CrossRef] [Green Version]
- Coller, H.A.; Grandori, C.; Tamayo, P.; Colbert, T.; Lander, E.S.; Eisenman, R.N.; Golub, T.R. Expression analysis with oligonucleotide microarrays reveals that MYC regulates genes involved in growth, cell cycle, signaling, and adhesion. Proc. Natl. Acad. Sci. USA 2000, 97, 3260–3265. [Google Scholar] [CrossRef] [Green Version]
- Okuyama, R.; Nguyen, B.C.; Talora, C.; Ogawa, E.; Tommasi di Vignano, A.; Lioumi, M.; Chiorino, G.; Tagami, H.; Woo, M.; Dotto, G.P. High commitment of embryonic keratinocytes to terminal differentiation through a Notch1-caspase 3 regulatory mechanism. Dev. Cell 2004, 6, 551–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, T.; Miyazaki, M.; Hakuno, F.; Takahashi, S.; Chida, K. PKCη promotes a proliferation to differentiation switch in keratinocytes via upregulation of p27Kip1 mRNA through suppression of JNK/c-Jun signaling under stress conditions. Cell Death Dis. 2011, 2, e157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Bellemare, J.; Roberge, C.J.; Bergeron, D.; Lopez-Vallé, C.A.; Roy, M.; Moulin, V.J. Epidermis promotes dermal fibrosis: Role in the pathogenesis of hypertrophic scars. J. Pathol. 2005, 206, 1–8. [Google Scholar] [CrossRef]
- Niessen, F.B.; Andriessen, M.P.; Schalkwijk, J.; Visser, L.; Timens, W. Keratinocyte-derived growth factors play a role in the formation of hypertrophic scars. J. Pathol. 2001, 194, 207–216. [Google Scholar] [CrossRef]
- Machesney, M.; Tidman, N.; Waseem, A.; Kirby, L.; Leigh, I. Activated keratinocytes in the epidermis of hypertrophic scars. Am. J. Pathol. 1998, 152, 1133–1141. [Google Scholar] [PubMed]
- Hakvoort, T.E.; Altun, V.; Ramrattan, R.S.; van der Kwast, T.H.; Benner, R.; van Zuijlen, P.P.; Vloemans, A.F.; Prens, E.P. Epidermal participation in post-burn hypertrophic scar development. Virchows Arch. 1999, 434, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Limandjaja, G.C.; van den Broek, L.J.; Waaijman, T.; van Veen, H.A.; Everts, V.; Monstrey, S.; Scheper, R.J.; Niessen, F.B.; Gibbs, S. Increased epidermal thickness and abnormal epidermal differentiation in keloid scars. Br. J. Dermatol. 2017, 176, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.S.; Joo, S.Y.; Cho, Y.S.; Park, J.H.; Ro, Y.M.; Kim, J.B.; Seo, C.H. Effect of extracorporeal shock wave therapy on keratinocytes derived from human hypertrophic scars. Sci. Rep. 2021, 11, 17296. [Google Scholar] [CrossRef]
- Bernard, B.A.; Asselineau, D.; Schaffar-Deshayes, L.; Darmon, M.Y. Abnormal sequence of expression of differentiation markers in psoriatic epidermis: Inversion of two steps in the differentiation program? J. Investig. Dermatol. 1988, 90, 801–805. [Google Scholar] [CrossRef] [Green Version]
- Boguniewicz, M.; Leung, D.Y. Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef] [Green Version]
- Martin-Medina, A.; Lehmann, M.; Burgy, O.; Hermann, S.; Baarsma, H.A.; Wagner, D.E.; De Santis, M.M.; Ciolek, F.; Hofer, T.P.; Frankenberger, M.; et al. Increased extracellular vesicles mediate WNT5A signaling in idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2018, 198, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Sun, X.; Liu, L.; Jiang, H.; Shen, Y.; Xu, X.; Li, J.; Zhang, G.; Huang, J.; Lin, Z.; et al. Exosomes and their therapeutic potentials of stem cells. Stem Cells Int. 2016, 2016, 7653489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.J.; Zhang, J.X.; Wang, R.L. Exosomes as mediators and biomarkers in fibrosis. Biomark. Med. 2020, 14, 697–712. [Google Scholar] [CrossRef]
- Zhang, X.; Yin, M.; Zhang, L.J. Keratin 6, 16 and 17-Critical Barrier Alarmin Molecules in Skin Wounds and Psoriasis. Cells 2019, 8, 807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, D.; Brash, D.E.; Grossman, D. Keratinocyte apoptosis in epidermal development and disease. J. Investig. Dermatol. 2006, 126, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Hockenbery, D.M.; Zutter, M.; Hickey, W.; Nahm, M.; Korsmeyer, S.J. BCL2 protein is topographically restricted in tissues characterized by apoptotic cell death. Proc. Natl. Acad. Sci. USA 1991, 88, 6961–6965. [Google Scholar] [CrossRef] [Green Version]
- Krajewski, S.; Krajewska, M.; Reed, J.C. Immunohistochemical analysis of in vivo patterns of Bak expression, a proapoptotic member of the Bcl-2 protein family. Cancer Res. 1996, 56, 2849–2855. [Google Scholar]
- Tomková, H.; Fujimoto, W.; Arata, J. Expression of the bcl-2 homologue bax in normal human skin, psoriasis vulgaris and non-melanoma skin cancers. Eur. J. Dermatol. 1998, 8, 256–260. [Google Scholar]
- Sayama, K.; Hanakawa, Y.; Shirakata, Y.; Yamasaki, K.; Sawada, Y.; Sun, L.; Yamanishi, K.; Ichijo, H.; Hashimoto, K. Apoptosis signal-regulating kinase 1 (ASK1) is an intracellular inducer of keratinocyte differentiation. J. Biol. Chem. 2001, 276, 999–1004. [Google Scholar] [CrossRef] [Green Version]
- Van de Craen, M.; Van Loo, G.; Pype, S.; Van Criekinge, W.; Van den Brande, I.; Molemans, F.; Fiers, W.; Declercq, W.; Vandenabeele, P. Identification of a new caspase homologue: Caspase-14. Cell Death Differ. 1998, 5, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Eckhart, L.; Declercq, W.; Ban, J.; Rendl, M.; Lengauer, B.; Mayer, C.; Lippens, S.; Vandenabeele, P.; Tschachler, E. Terminal differentiation of human keratinocytes and stratum corneum formation is associated with caspase-14 activation. J. Investig. Dermatol. 2000, 115, 1148–1151. [Google Scholar] [CrossRef] [Green Version]
- Hou, P.F.; Jiang, T.; Chen, F.; Shi, P.C.; Li, H.Q.; Bai, J.; Song, J. KIF4A facilitates cell proliferation via induction of p21-mediated cell cycle progression and promotes metastasis in colorectal cancer. Cell Death Dis. 2018, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Missero, C.; Calautti, E.; Eckner, R.; Chin, J.; Tsai, L.H.; Livingston, D.M.; Dotto, G.P. Involvement of the cell-cycle inhibitor Cip1/WAF1 and the E1A-associated p300 protein in terminal differentiation. Proc. Natl. Acad. Sci. USA 1995, 92, 5451–5455. [Google Scholar] [CrossRef] [Green Version]
- Di Cunto, F.; Topley, G.; Calautti, E.; Hsiao, J.; Ong, L.; Seth, P.K.; Dotto, G.P. Inhibitory function of p21Cip1/WAF1 in differentiation of primary mouse keratinocytes independent of cell cycle control. Science 1998, 280, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Hauser, P.J.; Agrawal, D.; Flanagan, M.; Pledger, W.J. The role of p27Kip1 in the in vitro differentiation of murine keratinocytes. Cell Growth Differ. 1997, 8, 203–211. [Google Scholar]
- Rangarajan, A.; Talora, C.; Okuyama, R.; Nicolas, M.; Mammucari, C.; Oh, H.; Aster, J.C.; Krishna, S.; Metzger, D.; Chambon, P.; et al. Notch signaling is a direct determinant of keratinocyte growth arrest and entry into differentiation. EMBO J. 2001, 20, 3427–3436. [Google Scholar] [CrossRef] [Green Version]
- Waikel, R.L.; Kawachi, Y.; Waikel, P.A.; Wang, X.J.; Roop, D.R. Deregulated expression of c-Myc depletes epidermal stem cells. Nat. Genet. 2001, 28, 165–168. [Google Scholar] [CrossRef]
- Descargues, P.; Sil, A.K.; Karin, M. IKKalpha, a critical regulator of epidermal differentiation and a suppressor of skin cancer. EMBO J. 2008, 27, 2639–2647. [Google Scholar] [CrossRef] [Green Version]
- Cabodi, S.; Calautti, E.; Talora, C.; Kuroki, T.; Stein, P.L.; Dotto, G.P. A PKC-eta/Fyn-dependent pathway leading to keratinocyte growth arrest and differentiation. Mol. Cell 2000, 6, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Shen, Y.; Mohanasundaram, P.; Lindström, M.; Ivaska, J.; Ny, T.; Eriksson, J.E. Vimentin coordinates fibroblast proliferation and keratinocyte differentiation in wound healing via TGF-β-Slug signaling. Proc. Natl. Acad. Sci. USA 2016, 113, E4320–E4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.S.; Cho, Y.S.; Joo, S.Y.; Mun, C.H.; Seo, C.H.; Kim, J.B. Wound healing potential of low temperature plasma in human primary epidermal keratinocytes. Tissue Eng. Regen. Med. 2019, 16, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.S.; Joo, S.Y.; Cho, Y.S.; Kim, J.B.; Seo, C.H. CPEB1 or CPEB4 knockdown suppresses the TAK1 and Smad signalings in THP-1 macrophage-like cells and dermal fibroblasts. Arch. Biochem. Biophys. 2020, 683, 108322. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|---|
| KRT1 | NM_006121.4 | TGGATGGTGCTTATATGAC | GACAACTCTGCTTGGTAG |
| KRT6A | NM_005554.4 | TGAAGAAGGATGTGGATG | ATCATACAAGGCTCTCAG |
| KRT10 | NM_000421.5 | GATTCTCAACCTAACAACTG | GCTACCTCATTCTCATACT |
| KRT14 | NM_000526 | GCTGAGATCAAAGACTACA | AGAAGGACATTGGCATTG |
| KRT16 | NM_005557.4 | CCTACTTCAAGACCATCG | CCTGGCATTGTCAATCTG |
| KRT17 | NM_000422.3 | ATCCTGCTGGATGTGAAGACGC | TCCACAATGGTACGCACCTGAC |
| MAP3K5 | NM_005923.4 | CGTGAGCACGCTCAGTTCTA | TTCCGAACCAATTCTTCCAG |
| NOTCH1 | NM_017617.5 | AGCCTCAACGGGTACAAG | TTGACACAAGGGTTGGATTC |
| CASP14 | NM_012114.3 | CCTGTTGTCACCTTGCTAT | GTCCTTGCCTCTGTCTTAC |
| BAX | NM_001291428.2 | CCTTTTGCTTCAGGGTTTCA | CCATGTTACTGTCCAGTTCG |
| BCL2 | NM_000633.3 | TGCGGCCTCTGTTTGATTT | AGGCATGTTGACTTCACTTGT |
| CDKN1A | NM_078467.3 | TAGGCGGTTGAATGAGAG | AAAGGAGAACACGGGATG |
| CDKN1B | NM_004064.5 | GCAGGAATAAGGAAGCGA | GGGAACCGTCTGAAACAT |
| PRKCH | NM_006255 | CGCCATCTTGAGACATCCTT | TTCTCGGGATTTGATTCTGG |
| VIM | NM_003380.5 | AAAGCGTGGCTGCCAAGAA | ACCTGTCTCCGGTACTCGTTTGA |
| CDH1 | NM_004360.5 | GCAGACCTTCCTCCCAATAC | TGGGTCGTTGTACTGAATGG |
| CDH2 | NM_001792.5 | CCACCTTA AAATCTGCAGGC | CCATGTTACTGTCCAGTTCG |
| MYC | NM_002467.6 | CCTGGTGCTCCATGAGGAGAC | CAGACTCTGACCTTTTGCCAGG |
| GAPDH | NM_ 002046.7 | CATGAGAAGTATGACAACAGCCT | AGTCCTTCCACGATACCAAAGT |
| Target | Host | Dilution | Company (Cat. No.) |
|---|---|---|---|
| CD 9 | Mouse | 1:250 | Invitrogen (10626D), Waltham, MA USA |
| CD 63 | Rabbit | 1:1000 | System Biosciences (EXOAB-CD63A-1), Palo Alto, CA, USA |
| CD 81 | Mouse | 1:250 | Invitrogen (10630D), Waltham, MA, USA |
| Ki-67 | Rabbit | 1:1000 | Abcam (ab16667), Cambridge, UK |
| Keratin 1 | Rabbit | 1:1000 | Abcam (ab93652), Cambridge, UK |
| Keratin 6 | Mouse | 1:2000 | Abcam (ab18586), Cambridge, UK |
| Keratin 10 | Rabbit | 1:500 | Santa Cruz Technology (sc-23877), Dallas, TX, USA |
| Keratin 14 | Rabbit | 1:2000 | Abcam (ab181595), Cambridge, UK |
| Keratin 16 | Rabbit | 1:2000 | Abcam (ab76416), Cambridge, UK |
| Keratin 17 | Rabbit | 1:2000 | Abcam (ab8068), Cambridge, UK |
| ASK 1 | Rabbit | 1:1000 | Abcam (ab45178), Cambridge, UK |
| Notch 1 | Rabbit | 1:1000 | Abcam (ab52627), Cambridge, UK |
| Caspase 14 | Rabbit | 1:1000 | Abcam (ab174847), Cambridge, UK |
| Bax | Rabbit | 1:1000 | Abcam (ab199677), Cambridge, UK |
| Bcl2 | Rabbit | 1:1000 | Abcam (ab196495), Cambridge, UK |
| P21 | Rabbit | 1:1000 | Abcam (ab109199), Cambridge, UK |
| P27 | Rabbit | 1:1000 | Abcam (ab32034), Cambridge, UK |
| PKCη | Rabbit | 1:1000 | GenScript (A00906), Piscataway, NJ, USA |
| Vimentin | Rabbit | 1:2000 | Abcam (ab92547), Cambridge, UK |
| E-cadherin | Rabbit | 1:1000 | Cell Signaling Technology (3195S), Danvers, MA, USA |
| N-cadherin | Mouse | 1:1000 | Thermo Fisher Scientific (33-3900), Waltham, MA, USA |
| MYC | Rabbit | 1:1000 | Cell Signaling Technology (9402), Danvers, MA, USA |
| β-actin | Rabbit | 1:2000 | Cell Signaling Technology (4967S), Danvers, MA, USA |
| β-actin | Mouse | 1:1000 | Santa Cruz Technology (sc-47778), Dallas, TX, USA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, H.S.; Joo, S.Y.; Lee, S.Y.; Cho, Y.S.; Kim, D.H.; Seo, C.H. Effect of Hypertrophic Scar Fibroblast-Derived Exosomes on Keratinocytes of Normal Human Skin. Int. J. Mol. Sci. 2023, 24, 6132. https://doi.org/10.3390/ijms24076132
Cui HS, Joo SY, Lee SY, Cho YS, Kim DH, Seo CH. Effect of Hypertrophic Scar Fibroblast-Derived Exosomes on Keratinocytes of Normal Human Skin. International Journal of Molecular Sciences. 2023; 24(7):6132. https://doi.org/10.3390/ijms24076132
Chicago/Turabian StyleCui, Hui Song, So Young Joo, Seung Yeol Lee, Yoon Soo Cho, Dong Hyun Kim, and Cheong Hoon Seo. 2023. "Effect of Hypertrophic Scar Fibroblast-Derived Exosomes on Keratinocytes of Normal Human Skin" International Journal of Molecular Sciences 24, no. 7: 6132. https://doi.org/10.3390/ijms24076132