
The Role of the NLRP3 Inflammasome in the Molecular and Biochemical Mechanisms of Cervical Ripening: A Comprehensive Review


1
Department of Perinatology, Gynecology and Gynecologic Oncology, Faculty of Health Sciences, Collegium Medicum in Bydgoszcz, Nicolaus Copernicus University, Łukasiewicza 1, 85-821 Bydgoszcz, Poland
2
Department of Obstetrics and Gynecology, St. Adalbert’s Hospital in Gdańsk, Copernicus Healthcare Entity, Jana Pawła II 50, 80-462 Gdańsk, Poland
*
Author to whom correspondence should be addressed.
Cells 2024, 13(7), 600; https://doi.org/10.3390/cells13070600
Submission received: 23 February 2024
/
Revised: 27 March 2024
/
Accepted: 27 March 2024
/
Published: 29 March 2024
(This article belongs to the Special Issue Signaling Pathways in Pregnancy)
Abstract
:The uterine cervix is one of the key factors involved in ensuring a proper track of gestation and labor. At the end of the gestational period, the cervix undergoes extensive changes, which can be summarized as a transformation from a non-favorable cervix to one that is soft and prone to dilation. During a process called cervical ripening, fundamental remodeling of the cervical extracellular matrix (ECM) occurs. The cervical ripening process is a derivative of many interlocking and mutually driving biochemical and molecular pathways under the strict control of mediators such as inflammatory cytokines, nitric oxide, prostaglandins, and reactive oxygen species. A thorough understanding of all these pathways and learning about possible triggering factors will allow us to develop new, better treatment algorithms and therapeutic goals that could protect women from both dysfunctional childbirth and premature birth. This review aims to present the possible role of the NLRP3 inflammasome in the cervical ripening process, emphasizing possible mechanisms of action and regulatory factors.
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Seemingly insignificant, the uterine cervix is the main factor ensuring the smooth course of both pregnancy and labor. During a normal pregnancy, the cervix is responsible for providing a physical and immunobiological barrier, ensuring unrestricted intrauterine fetal maturing. In the final stage of term gestation (or during premature delivery), profound changes occur in the cervical tissue, leading to extensive changes in cervical tissue composition. Macroscopically, these changes are manifested by transformation from rigid and tender to soft and prone to dilation. This change in the structure of the cervical tissue allows it to dilate properly during uterine contractions. Macroscopic changes in the cervix’s consistency result from several interrelated biochemical and molecular pathways leading to a total rearrangement of the extracellular matrix of the cervical tissue stroma. The set of these numerous intertwining metabolic pathways can be generally summarized as the cervical ripening process [1]. Cervical tissue remodeling is as prerequisite for proper parturition. As a result of the above processes, the cervix increases its softness and susceptibility, which allows labor to occur.
Despite the countless studies on cervical tissue metabolism, the whole process of cervical remodeling is poorly understood. The complex process of cervical ripening comprises a vast array of well-orchestrated biochemical pathways in which cooperation is required for the proper course of cervical remodeling. Numerous regulatory factors are known to have a significant impact on this process. These factors mainly include cytokines, metalloproteinases, prostaglandins, hormones, transcription factors, reactive oxygen species, etc. [2]. However, more and more attention has recently been assigned to the role of the local inflammatory response in cervical ripening. Inflammatory events occurring in cervical stroma include invasion of inflammatory cells (such as neutrophils and macrophages) and increased levels of inflammatory cytokines [2]. At the same time, these processes are strictly regulated both at the cellular and molecular levels [2,3].
Inflammasomes are a class of multimeric protein complexes that mediate the activation of potent inflammatory factors [4]. They are integral components of the innate immune response and actively participate in the development of the inflammatory cascade through their activation by inflammatory factors or stressors. Nucleotide-binding domain, leucine-rich-containing family, pyrin domain-containing-3 (NLRP3), which is one of the best-known and studied inflammasomes, greatly contributes to the inflammatory process by mediating the secretion of interleukin-1 (IL-1), which is a pivotal pro-inflammatory cytokine [5]. Numerous studies indicate the active involvement of the NLRP3 inflammasome in the development of the inflammatory response during pregnancy-associated complications [6]. However, whether the NLRP3 inflammasome is implicated in the cervical tissue remodeling process is largely unknown and poorly understood. Additionally, it is not fully understood what impact the NLRP3 inflammasome may have on cervical tissue biology.
In this review, we would like to discuss the role of the NLRP3 inflammasome in the process of cervical ripening, taking into account possible mechanisms of action and emphasizing its possible regulatory factors.
2. Uterine Cervix Biology
The uterine cervix can be divided into vaginal and supravaginal portions [7]. The cervical canal is a center structure of the cervix and is lined with a cylindrical epithelium with a single layer of cells producing significant amounts of mucus, containing ions, enzymes, mucin glycoproteins, and plasma proteins [7]. As gestation progresses, cervical epithelial cells proliferate significantly, leading to increased production of cervical mucus. Due to specific mucus formation, it is possible to provide an immunological barrier that ensures unimpeded fetal intrauterine development [7,8].
The uterine cervix is composed, in particular, of ground substance, connective tissue, and smooth muscle with the addition of cellular elements. The cervical structure is infiltrated by vast numbers of blood vessels, nerves, and lymphatic vessels. The cellular compartment of the cervix is composed mainly of fibroblasts, mast cells, and wandering cells [9].
The smooth muscle content of the cervical tissue accounts for approximately 15% of the dry mass of the cervix [9]. The distribution of muscle fibers along the cervix is extremely interesting. The area of the external cervical os is intertwined with muscle fibers, which constitute approximately 10% of the cervix, and these fibers are loosely dispersed in space. In turn, in the area of the internal os, muscle fibers are present in much larger numbers [9,10]. Additionally, fibers located around the internal cervical os undergo much more advanced spatial organization and form a characteristic sphincter-like pattern [10]. This specific spatial organization of the internal cervical os may suggest the inherence of the sphincter-like fiber system.
Ground substance, also known as the extracellular matrix (ECM), is the basic building block of the cervix [1]. The main components of the ECM are collagen types I and III, with a small percentage of collagen IV [1,11]. Collagen production is provided by fibroblasts, which actively participate in the proper metabolism of cervical tissue [11]. Interestingly, in the peripartum period, a significant decrease in the expression of type I collagen mRNA in cervical tissue can be observed [12]. The collagen fibers of the cervical stroma are connected to each other by covalent bonds [11]. The formation of covalent bonds enables the creation of a durable (degradation-resistant) three-dimensional structure, which ensures the stiffness of the cervix during pregnancy [11,13]. The cervical stroma is composed of glycosaminoglycans (GAGs), which contain vast amounts of sulfate groups [14]. The presence of a large amount of sulfate residues ensures high hydrophilic properties of the GAG molecule. The most important GAGs that build the cervical ECM include dermatan sulfate, chondroitin sulfate, heparan sulfate, and hyaluronan (HA) [14]. GAGs have the ability to covalently bond the protein core, leading to the formation of proteoglycans. Proteoglycans can subsequently bind free collagen fibers via their free anionic residue [13,14,15]. There are three main types of proteoglycans that make up the cervical ground substance: chondroitin (PG-L), decorin (PG-S2), and biglycan (PG-S1) [16]. The formation of such a three-dimensional structure composed of proteoglycans and collagen fibers enables the creation of a compact, stiff structure that is resistant to proteolytic degradation [11,12,13,14,15,16,17,18,19]. Taking the above into consideration, it seems that qualitative changes in proteoglycans or collagen arrangement may have profound effects on cervical tissue mechanics.
Elastin fibers also contribute to building the stroma of the cervix [9,20]. They consist of elastin polymers, which are cross-linked into more advanced spatial structures known as microfibrils [21]. In the non-pregnant stage, microfibrils show a long, organized, and well-organized structure. However, during gestation progression, elastic fibers’ concentration gradually decreases, and their structure becomes less organized and more randomly scattered in cervical tissue [21,22].
As mentioned earlier, during the perinatal period, the cervical tissue undergoes fundamental structural changes characterized as cervical ripening. These structural alterations can generally be described as a spatial remodeling of the cervical ECM. The descending effect of these changes is a significant loosening of the structure of the previously tightly packed cervix and an increase in the softness and compliance of the vaginal portion of the uterine cervix [23,24]. From a histological point of view, the main changes occurring in the cervix are the degradation of collagen and elastin fibers (accompanied by a decrease in total collagen concentration), increased water inflow (causing an increase in tissue hydration), increased HA synthesis, an increase in cervical cells’ apoptosis, and the occurrence of a local inflammatory response [9,11]. The whole process combines a vast range of interwoven (and interdependent) biochemical and molecular processes that are strictly controlled by regulatory factors.
During the gestational period, the synthesis of HA gradually increases, with the greatest increase in the perinatal period. Moreover, the transcription of genes encoding the hyaluronan synthase (HAS) enzyme increases throughout pregnancy—mainly HAS2 isoform [19,20]. In the perinatal period, HA remains the dominant GAG in cervical tissue [20]. An increased HA concentration in the cervical stroma, due to its hydrophilic properties, causes a significantly increased inflow of water into the ECM. As a result, an increased dispersion of collagen and elastin fibers in the cervix occurs during its remodeling [11,19,20,21,22,23].
The change that is most pronounced during cervical ripening is undoubtedly the decrease in total collagen concentration. This is achieved mostly by reducing the expression of collagen assembling genes and by increasing the enzymatic breakdown of collagen fibers [18,24,25,26,27,28].
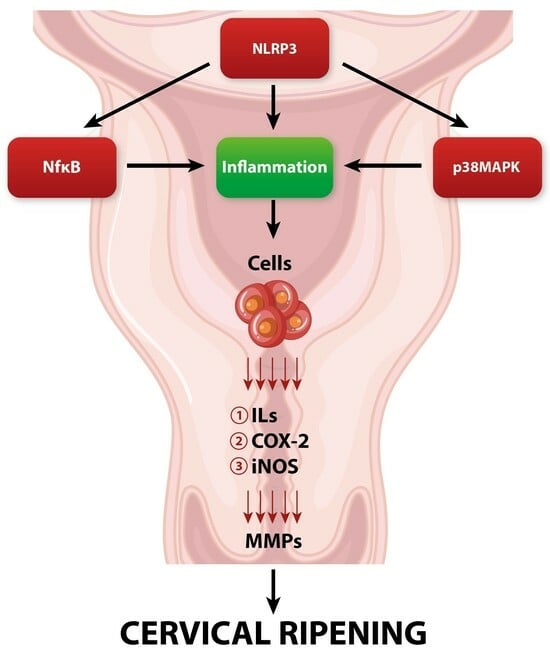
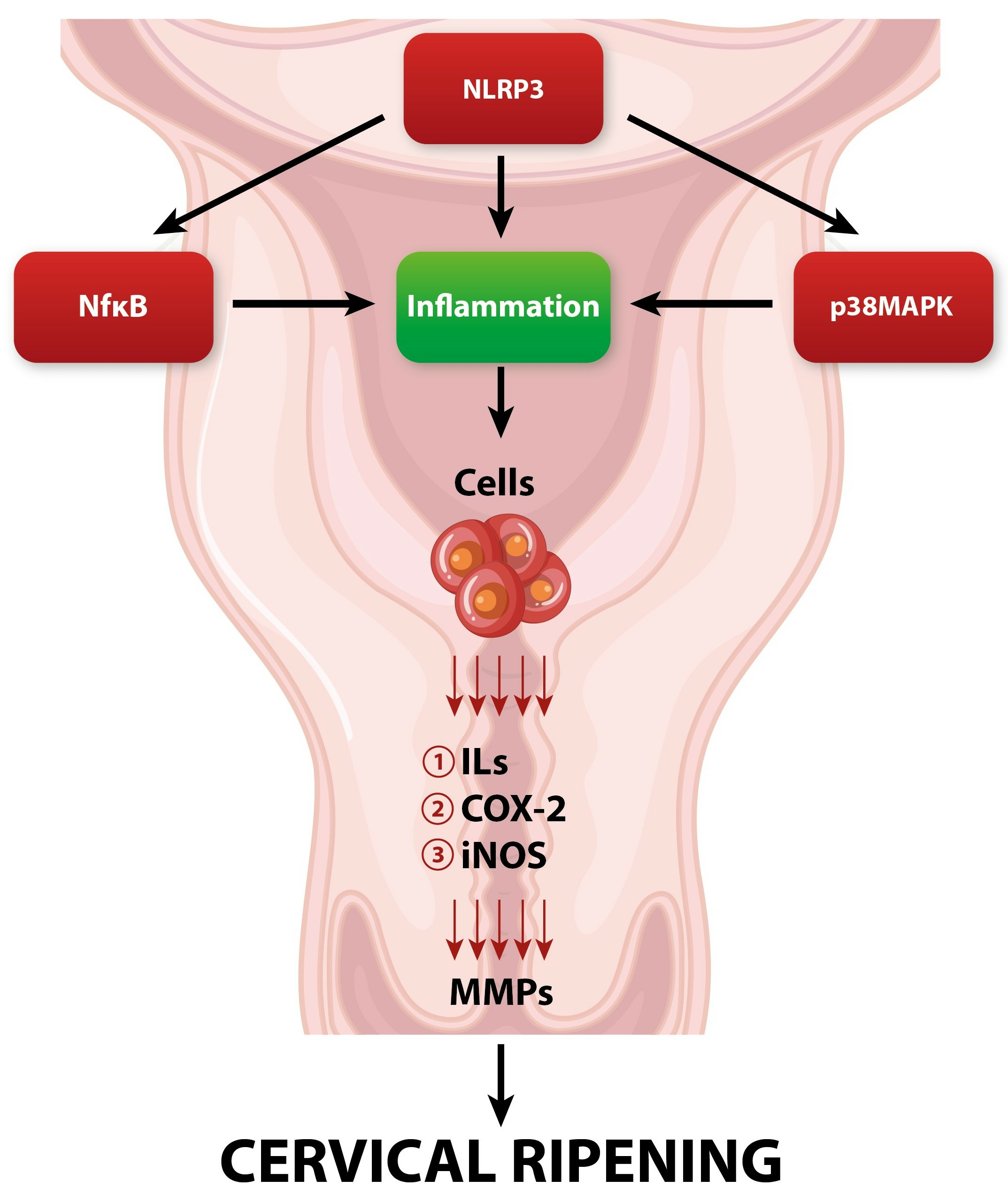
Enzymatic cleavage of collagen fibers is maintained by matrix metalloproteinases (MMPs) secreted mainly by fibroblasts, macrophages, neutrophils, and stromal cells [29]. MMPs are the main executors of the actual remodeling of the cervical ECM during its ripening [29,30]. In the perinatal period, there is a gradual increase in the activity and concentration of MMPs in the cervical tissue [31]. MMPs are synthesized and secreted in the form of prepro-enzymes (zymogens), which achieve their active form after enzymatic cleavage [32]. Activated MMPs can actively digest covalent bonds between collagen fibers, causing a change in ECM spatial organization—from tightly packed large multimers of collagen fibrils to smaller, loosely scattered bundles of fibers [33]. In turn, collagen fibers undergo spatial reorganization, leading to the significant softening of the cervix. Moreover, in addition to collagen fibers’ enzymatic breakdown, MMPs also have the ability to enzymatically cleave other ECM elements such as PG-S2, PG-L, and fibronectin [11]. The secretion and enzymatic activity of MMPs are closely regulated by other factors involved in pregnancy and parturition [34]. The expression of MMPs is enhanced by pro-inflammatory cytokines, nitric oxide (NO), prostaglandins (PGs), and growth factors [34,35]. In turn, their activity is suppressed by progesterone, retinoic acid, glucocorticoids, or specific tissue inhibitors of metalloproteinases (TIMPs) [36,37]. The cervical ripening process is under the strict influence of a variety of regulatory factors, which we will discuss in further sections of this review (Figure 1).
3. NLRP3 Inflammasome Biology
Inflammasomes are large cytoplasmic protein multimers that assemble in response to the detection of stress-associated or infectious stimuli [38]. They are pivotal for immune defenses against a variety of pathogens. Inflammasomes are involved in the immunological response against viral and bacterial infections [39,40]. Additionally, inflammasomes are crucial in maintaining cell homeostasis during stress-related events or environmental irritants [41]. Pattern-recognition receptors (PRRs) are a family of receptors that have the ability to be activated upon exposure to exogenous stimuli such as pathogens or a variety of endogenous factors [5]. PRRs recognize pathogen components known as pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs), which are generated endogenously by cells subjected to a damaging factor [42]. Subsequently, those reactions trigger inflammatory response pathways in the cell environment. PRRs, apart from being present in human cells, can actively participate in inflammasome formation. Several PRRs have been shown to have the ability to form inflammasome multimers: the leucine-rich (LRR) repeat-containing protein family (NLRP1, NLRP3, etc.), pyrin, or absent-in-melanoma 2 protein (AIM 2) [43]. The names of the resulting inflammasomes arise from the names of the PRR sensor domains that constitute the inflammasome multimers during the inflammatory response, e.g., the NLRP3 inflammasome [5,42,43].
3.1. NLRP3 Inflammasome Structure
A typical inflammasome consists of a sensor protein (PRR), the adaptor apoptosis-associated speck-like protein (ASC) containing a caspase recruiting domain, and pro-inflammatory pro-caspase-1, which is responsible for further catalytic functions [44]. The formation of the inflammasome causes the activation of caspase-1, which subsequently cleaves the pro-inflammatory cytokines pro-interleukin-1β (pro-IL-1β) and pro-IL-18 into their mature, active forms—interleukin-1β (IL-1β) and interleukin-18 (IL-18) [45,46]. Subsequently, enzymatically cleaved interleukins participate in the inflammatory response. Apart from interleukins’ enzymatic activation, caspase-1 has the ability to trigger gasdermin D (GSDMD) activation, which participates in cell membrane pore formation, thereby activating the inflammatory form of cell death—pyroptosis [47]. The aim of pyroptosis is to remove potential pathogenic factors from the cell, allowing for their degradation by immunological factors. Additionally, as a result of pyroptosis, DAMPs and activated interleukins are removed from the cell, which may cause further activation of the inflammatory reaction in nearby cells [47,48,49,50].
The NLRP3 inflammasome is by far the most studied and well-known inflammasome, which is critical for proper host immune defense systems against pathogens such as bacterial, fungal, and viral infections. Additionally, NLRP3 has been linked with the pathogenesis of several diseases, such as Alzheimer’s disease or atherosclerosis [51]. NLRP3 is a tripartite molecule of the leucine-rich repeat (NLR) family, which consists of a central nucleotide-binding and oligomerization domain (NACHT or NOD), amino-terminal pyrin domain (PYD), and C-terminal leucine-rich repeat domain (LRR) [5,52]. The NLRP3 PYD domain is required for proper interaction with the PYD domain of the ASC protein. The NACHT domain has ATPase activity, which is responsible for the conformational change and oligomerization of the NLRP3 inflammasome [53]. The LRR domain has a receptor function and is responsible for external signal transduction [54]. The ASC protein of the forming inflammasome has a PYD domain (identical to the PYD domain of NLRP3) and a caspase activation and recruitment domain (CARD) that bonds to the corresponding CARD domain of caspase-1. Then, as a result of ATPase activation of the NACHT domain, oligomerization occurs, and a functional inflammasome is formed. Upon response to the stimuli, NLRP3 oligomerizes (leading to the spatial conformation of the multimer alteration) through NACHT domain interaction. Then, the ASC protein is recruited via PYD–PYD domain bonding. Assembled ASC complex recruits enable auto-cleavage and further activation of the caspase-1 enzyme [5,52,53,54].
3.2. NLRP3 Inflammasome Activation and Regulation
There are two possible routes to the formation and activation of the NLRP3 inflammasome—the canonical and non-canonical inflammasome assembly pathways [55]. The canonical pathway requires two signals for inflammasome activation—the priming and activation signals [56]. The priming signal is initiated by various PAMPs, IL-1β, Toll-like receptor (TLR) ligands, or tumor necrosis factor (TNF) [5]. The binding of specific PAMPs to their cell receptors induces the activation of nuclear factor-kappa-beta (NF-κB) with its subsequent translocation to the nucleus, where it enhances the transcription of genes encoding the NLRP3 protein (which belongs to the PRR group), pro- IL-1β, pro-IL-18, and gesdermin D [57,58,59]. Additional factors such as caspase-8 and FAS-associated death domain (FADD) are required for proper NF-κB nuclear translocation [60]. Apart from the correct priming signal, additional regulatory factors are required for proper NF-κB activation and translocation. Caspase-8 and FADD can interact with the IκB kinase (IKK) complex, which inhibits the transcriptional activity of NF-κB. Those regulating factors, by binding to IKK, cause the disinhibition of the NF-κB molecule, enabling its translocation to the nucleus, where it enhances NLRP3 and pro-IL-1 genes’ transcription [60]. The priming signal is necessary for NLRP3 to become more responsive to an activation signal. After the priming signal, the second stage occurs—the activation signal, which is triggered by a variety of PAMPs and DAMPs. The second signal step consists of NLRP3 oligomerization, ASC clustering, and caspase-1 bonding, leading to the full NLRP3 inflammasome assembly [61]. After the PYD domains of NLRP3 and ASC interact, the CARDS domain of ASC gains the ability to bond the CARD domain of pro-caspase-1. At the same time, the NACTH domain induces the oligomerization of the forming NLRP3 inflammasome. As a result, a functioning, mature NLRP3 inflammasome assembles [62]. Subsequently, after enzymatic self-cleavage, caspase-1 is released, leading to the activation and secretion of IL-18 and IL-1β [5,57,58,59,60]. Additionally, mature caspase-1 induces pyroptosis via gesdermin D activation. Caspase-1 cleaves GSDMD into N-term GDMD, which forms pores in the cell membrane, leading to the subsequent cytokines’ release from the cell [63,64,65]. Released cytokines can act as other DAMPs, leading to the activation and formation of new inflammasomes in neighboring cells. We believe that this two-phase pathway of inflammasome assembly and activation seems to be explained by macrophages’ biology, which are the main cells involved in the inflammatory reaction. The inheritance of NLPR3 activators is an insufficient signal for the overall assembly of the mature inflammasome [61]. Activated NF-κB enhances the NLRP3 expression, which is believed to be at levels inadequate to trigger inflammasome activation under physiological conditions [61]. However, while the primary signal has an effect on NLRP3 and IL-1β concentrations, it appears to have no effect on IL-18 and pro-caspase-1 concentrations [58]. This may suggest that the activation (second) signal is crucial for proper assembly and full activation of a functional NLRP3 inflammasome.
Various factors, mainly PAMPs and DAMPs, can induce an activation signal (second-stage signal) for NLRP3 inflammasome assembly. These factors do not appear to influence NLRP3 directly but through several common, overlapping biochemical pathways that lead to the activation of the NLRP3 inflammasome [5,11,66]. The activation of the NLRP3 inflammasome is stimulated by DAMPs, also known as alarmins, such as IL-1β, ATP, reactive oxygen species (ROS), high mobility group box-1 (HMGB1), and uric acid crystals [66]. PAMPs, meanwhile, refer to molecular fragments of pathogens such as bacterial RNA, bacterial DNA, fungi, and viral DNA [66,67,68,69].
Multiple molecular and biochemical cellular pathways that are activated by NLRP3 stimuli, including ionic flux, and ROS production have been shown to lead to NLRP3 inflammasome formation. Moreover, those substances can also act as alarmins and enhance the further NLRP3 priming signal, leading to the formation of inflammasomes in neighboring cells [5,62].
When a specific alarm signal reaches the cell, the primary signal is activated, and several biochemical and molecular pathways are triggered, inducing a second inflammasome activation signal. The NLRP3 stimuli can lead to an alteration in the ion potential of cells. These changes are mainly expressed by changes in the concentrations of calcium, potassium, chloride, and sodium ions [70]. When a damage signal occurs (e.g., PAMPs or DAMPs), the ATP-dependent ion channel P2RX7 is activated, which causes the efflux of K+ from the cell [70,71]. A decrease in the concentration of K+ in the cytoplasm causes the strengthening of the interaction between the never-in-mitosis gene A-related kinase 7 (NEK7) and NLRP3, leading to the activation of the NLRP3 inflammasome [62,72]. NEK7 belongs to the NIMA-related kinase (NEK proteins) serine/threonine kinase family and is indispensable for the NLRP3 inflammasome’s proper assembly [73]. The LRR domain of NLRP3 can bond with the catalytic domain of NEK7 in a kinase-dependent manner [74]. It seems that NEK7 is a crucial factor required for proper inflammasome activation. Recent studies report that caspase-1 and IL-1 secretion were suppressed in the absence of NEK7 [72]. Moreover, cells lacking NEK7 were unable to conduct proper ASC oligomerization [75]. This may suggest that despite the presence of a trigger signal, the lack of NEK7 activity results in the inability to properly assemble and activate the NLRP3 inflammasome. We believe that targeted anti-NEK7 treatment could prove effective in suppressing inflammasome assembly in a number of pregnancy-associated complications. However, this interesting topic requires further, in-depth research on the interrelationships between NEK7 and other factors regulating the inflammasome activation process.
At the same time, excess potassium ions cause damage to mitochondria, which increases ROS synthesis [76]. Additionally, the previously activated P2RX7 channel also enables the transport of calcium ions into the cell. The increase in Ca2+ in the cell also occurs through the activation of CASR receptors [77]. Ca2+ causes the activation of phospholipase C (PLC), which metabolizes phosphatidylinositol 4,5-bisphosphate (PIP2) to diacylglycerol (DAG) and 1,4,5-triphosphate (IP3). As a result, IP3, via binding to the endoplasmic reticulum (ER) membrane, causes the release of an additional portion of Ca2+ from the ER into the cytosol [78]. Excess calcium ions can cause direct formation of the NLPR3-ASC complex [78]. Additionally, overproduction of Ca2+ may lead to increased ROS production in the mitochondria [77]. Finally, a rapid change in the cell’s ionic potential may cause damage to the mitochondrial membrane, which will additionally translate into increased ROS production and the release of cardiolipin into the cytoplasm, which is an activator of apoptosis [61,69]. Additionally, the change in ionic potential also leads to damage to the lysosomal membrane, causing the release of cathepsin B into the cytoplasm, which has lysosomal protease activity [57]. Cathepsin B (as well as K+ and Ca2+) may directly activate specific kinases such as Janus kinases (JAKs) and Tat-associated kinases (TAKs), which can directly enhance the activation of the NLRP3 inflammasome [77,78,79,80,81,82].
Studies have revealed that knockdown of membrane chloride channels CLIC1 and CLIC4 impairs the transcription and secretion of pro-IL-1β, suggesting the active involvement of chlorine ions in the action of the inflammasome assembly [83,84]. CLICs are transmembrane channels responsible for Cl− efflux [84]. Overproduction of ROS by mitochondria can induce the translocation of CLICs to cell membranes, which will lead to the removal of chloride ions from cells [85]. A decrease in the Cl− concentration in cells leads to a direct increase in NEK7-NLRP3 interaction and subsequent assembly of the inflammasome [82,83]. Moreover, the decrease in Cl− concentration induces oligomerization of ASC domains, which stimulates inflammasome assembly. However, this process does not occur in the absence of K+ efflux [82,83,84,85,86,87]. This may suggest that changes in ion concentration are interdependent and interact with each other in the process of inflammasome activation and assembly. However, this topic requires further research.
The non-canonical inflammasome activation pathway refers to the caspase-11-dependent inflammasome assembly [61]. This specific pathway occurs without priming and activating signals, as in the canonical pathway. This activation pathway is mainly associated with bacterial infections and contributes to the development of septic shock [88]. During the development of a bacterial infection, bacterial lipopolysaccharide (LPS) penetrates the cytoplasm, where it directly activates caspase-11 [89,90,91,92]. Caspase-11 then enzymatically cleaves GSDMD into N-GSDMD and C-GSDMD, which induces pyroptosis [93]. Additionally, active caspase-11 activates the release of potassium from the cell, which is a signal for the activation of the NLPR3 inflammasome, leading to the secretion of IL-1β [93,94,95]. Furthermore, cleaved N-GSDMD creates cell membrane pores during pyroptosis, facilitating IL-1 secretion [95,96,97].
In addition to the canonical and non-canonical activation pathways, there is also an alternative pathway for NLRP3 inflammasome activation [56]. This pathway requires only one activation signal (including LPS), which activates caspase-8. Active caspase-8 then directly activates the NLRP3 inflammasome [88,89,90,91,92,93].
4. Inflammatory Response and NLRP3 Inflammasome during Cervical Remodeling
As mentioned before, molecular and biochemical pathways occurring during cervical ripening greatly correlate with the local inflammatory reaction. The vast number of factors involved in triggering and maintaining the inflammatory response are also powerful and potent cervical ripening mediators [34].
4.1. Inflammation during Cervical Ripening and Its Regulatory Factors
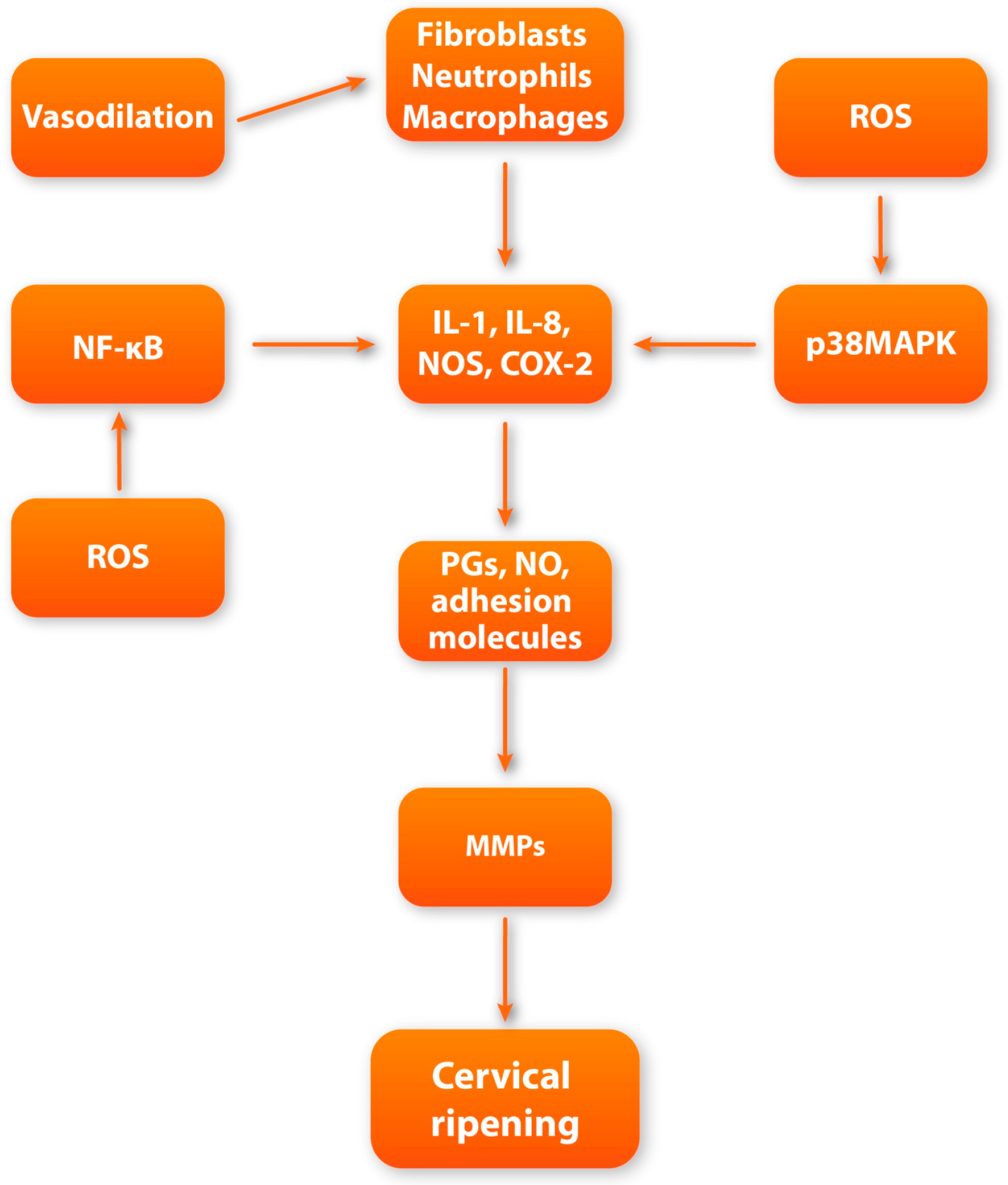
Local vasodilatation and escalation of vascular permeability occur during cervical ripening [98]. The downstream effect of increased vascular permeability is the increased influx of leukocytes and water into the cervical ECM [98]. Subsequently, significant swelling of the cervical stroma can be observed. Referring to the research, the end of gestation is accompanied by a constant increase in the concentration of inflammatory cells both in the cervix and in other maternal–fetal compartments such as fetal membranes or the myometrium [99]. Among the inflammatory cells that infiltrate the cervical tissue at the end of pregnancy, the dominant cells are macrophages and neutrophils [100]. The rapid increase in granulocytes’ concentration is possibly due to the increased expression of chemokines and adhesion molecules. In the perinatal period, there is a significant increase in the expression of CXC chemokine ligand 8 (CXCL8) in cervical tissue, which acts as a strong neutrophilic chemoattractant [101]. Macrophages and neutrophils are among the most important factors involved in cervical ripening through their ability to secrete large amounts of MMPs, pro-inflammatory cytokines, prostaglandins, and ROS [11]. Due to their degranulation, the concentration of factors such as IL-1, IL-6, IL-8, nitric oxide (NO), prostaglandins (PGs), tumor necrosis factor (TNF), and adhesion molecules significantly increases [99]. All of the above molecules are strongly involved in triggering local inflammatory reactions and maintaining the proper cervical ripening process.
IL-1 is one of the key players and regulators of cervical ripening [102]. Studies suggest that intravaginal application of interleukin-1 leads to a great increase in cervical compliance, which is accompanied by an increased concentration of neutrophils and increased activity of neutrophilic proteases [103]. IL-1 appears to affect cervical ripening at multiple levels. First, IL-1 directly increases the expression of MMPs while inhibiting the activity of TIMPs, which leads to increased collagenolytic activity in cervical tissue [11,104,105]. Moreover, IL-1 can enhance the synthesis of other pro-inflammatory cytokines, such as TNF, IL-6, and IL-8 [106]. Finally, it has a high affinity for prostaglandin metabolism, causing inhibition of prostaglandin dehydrogenase (PGDH) and stimulating cyclooxygenase-2 (COX-2) expression. As a result, due to the action of IL-1, metabolism is inhibited, and the synthesis of prostaglandins in the cervical stroma is increased, resulting in a great increase in PG concentration [107,108].
Prostaglandins appear to influence the cervix through direct and indirect effects on the ECM. PGs increase the overall water content in the ECM and also stimulate fibroblasts to synthesize GAG. PGs also have the ability to regulate the activity of inflammatory cells [109,110]. By stimulating the secretion of endothelial adhesion molecule (ICAM-1), they enable adhesion with subsequent infiltration of leukocytes into the cervical tissue [109]. Additionally, prostaglandins inhibit the secretion of secretory leukocyte protease inhibitor (SLPI), which acts as a potent neutrophil inhibitor [111]. However, their most important impact in the context of cervical remodeling is the direct stimulation of the secretion and activity of MMPs, which leads to the enzymatic degradation of collagen fibers [112,113].
Another crucial regulator of the proper cervical ripening process, synthesized by macrophages and neutrophils, is IL-8. According to research, IL-8 synthesis increases significantly in cervical tissue at term [11,114]. IL-8 mostly influences cervical ripening by increasing the secretion of MMPs [115]. IL-8 not only enhances ECM remodeling but also greatly influences the development of a local inflammatory reaction. By directly stimulating vascular permeability and chemotactic activity, IL-8 leads to an increased influx and activation of neutrophils at the site of inflammation [116,117,118].
Another important factor that not only triggers but also facilitates the proper course of a vast array of biochemical pathways of cervical ripening is nitric oxide (NO) [119]. Nitric oxide is produced in a reaction mediated by nitric oxide synthase (NOS). The dominant form of NOS involved in cervical metabolic pathways is inducible nitric oxide synthase (iNOS). The main sources of NO in cervical tissue are macrophages and neutrophils that have the ability to express iNOS [11,120,121]. There is ample evidence regarding the direct involvement of NO in cervical ripening. In the research conducted, the local administration of NO donors led to the effective triggering of cervical ripening [122]. Meanwhile, the administration of NOS inhibitors led to the inhibition and delay of cervical ripening pathways in cervical tissue [123]. These data may suggest that nitric oxide is a potent inducer of cervical remodeling during the perinatal period. For instance, nitric oxide greatly affects the cervical stromal composition. Beyond this, the impact of NO on cervical tissue is mostly manifested by enhancing the activity of MMPs, which cleaves collagen cross-links [124]. The indirect effect is reflected by promoting the influx of leukocytes into the cervical ECM by inducing local vasodilation with additional stimulation of IL-8 secretion [125,126]. Moreover, NO can directly stimulate COX-2 expression in cervical cells, leading to an increase in the secretion of PGs [127,128,129].
Tumor necrosis factor alpha (TNF-α) belongs to the inflammatory cytokine group and resembles the pleiotropic effects of a variety of cells [130]. It is a homotrimeric molecule actively involved in the inflammatory response, which is generated mostly by activated macrophages [130]. Similarly to other pro-inflammatory factors, an increase in TNF secretion in the cervix is notable in the perinatal period [11]. The role of TNF-α in the local inflammatory reaction is mainly to stimulate the secretion of other pro-inflammatory cytokines such as IL-1, IL-6, and IL-8 [131]. Additionally, TNF-α is a known powerful stimulator of the activity of phospholipase A2, which is the main enzyme involved in the synthesis of PGs [130]. Moreover, TNF-α can induce the release of neutrophil granules, leading to an increase in the concentration of neutrophil proteases. Finally, TNF-α can also activate NF-κB, leading to increased expression of other pro-inflammatory factors involved in cervical remodeling [130,131,132]. Taking the above into account, it seems that TNF-α can actively participate in the alteration of the cervical ECM composition.
In addition to the above-mentioned molecules that directly participate in cervical ripening, it is also worth noting the factors that can regulate and connect numerous biochemical pathways that contribute to cervical ripening, such as nuclear factor kappa-B (NF-κB). NF-κB is a group of transcription factors that consist of molecules such as p50, cRel, RelA/p65, p52, and RelB [59]. NF-κB remains under the heavy influence of inflammatory factors, glucocorticoids, and progesterone, which are crucial factors involved in cervical ripening [133]. Activation of NF-κB results in its translocation to the nucleus, where it subsequently enhances specific inflammatory genes’ transcription. In turn, the expression of inflammatory reagents such as cytokines (IL-6, IL-8, and IL-1), iNOS, and PGs is enhanced. [133,134,135]. Subsequently, increased transcription of genes encoding the mentioned pro-inflammatory factors increases their expression in the cervical tissue, which leads to enhancement in remodeling during cervical maturation. Moreover, NF-κB activity is enhanced by ROS (formed by NADPH neutrophilic oxidase), PGs, and nitric oxide [133,134,135,136,137,138,139]. It seems that in addition to stimulating the expression of pro-inflammatory factors by NF-κB, these factors have the ability to additionally stimulate the activity of NF-κB. These interrelationships may suggest the presence of a regulatory circuit in which NF-κB is the key player. We believe that the increase in the concentration of pro-inflammatory mediators during the local inflammatory response in cervical tissue, by positively influencing NF-κB activity, leads to the maintenance of permanent NF-κB nuclear translocation, leading to the unimpeded transcription of NF-κB-dependent inflammatory genes [140].
Mitogen-activated protein kinases (MAPKs) are proline-directed threonine and serine protein kinases, which are pivotal players in the macrophage-mediated inflammatory response [141]. The p38 proteins are a class of MAPKs that are greatly expressed in macrophages. p38MAPKs can be classified into four subtypes: α (MAPK14), β (MAPK11), γ (MAPK12/ERK6), and δ (MAPK13/SAPK4), with the dominance of the α subtype [141,142]. They are activated by a vast array of factors, including ROS, cytokines, pathogens, growth factors, and estrogen [141].
The most well-known MAPK cascade consists of extracellular-signal-regulated (ERK) kinases MEK1/2 and ERK 1/2 (p42/p44) [141]. Phosphorylation of MAPKs leads to their activation via ERK kinases’ phosphorylation. Once activated, MAPKs predominantly regulate gene expression via phosphorylation of downstream transcription factors, which include the pro-inflammatory transcription factors activator protein (AP)-1 and NF-κB, leading to an increase in the expression of inflammation-related factors [141,142]. The evidence strongly suggests that apart from participating in the systemic macrophage-mediated inflammatory response, p38 is also involved in cervical ripening inflammatory pathways [143,144]. The involvement of p38MAPK in these pathways may be manifested through the increased expression of COX-2 and pro-inflammatory cytokines (such as IL-1, IL-8, and IL-6) [145,146,147,148,149]. Moreover, p38 can stimulate the secretion of endothelial vascular cell adhesion molecule-1 (VCAM-1), which is responsible for leukocyte inflow during the inflammatory response [149]. In addition, active p38MAPK may directly enhance the synthesis of MMPs (especially MMP-9), which are key players involved in the enzymatic cleavage of collagen fibers in the stroma of the cervix [149,150,151,152,153]. Taking the above into account, it seems that p38MAPK is an important regulatory factor that integrates several metabolic pathways and actively participates in triggering the inflammatory reaction during cervical ripening.
4.2. NLRP3 Inflammasome in the Cervical Inflammatory Response
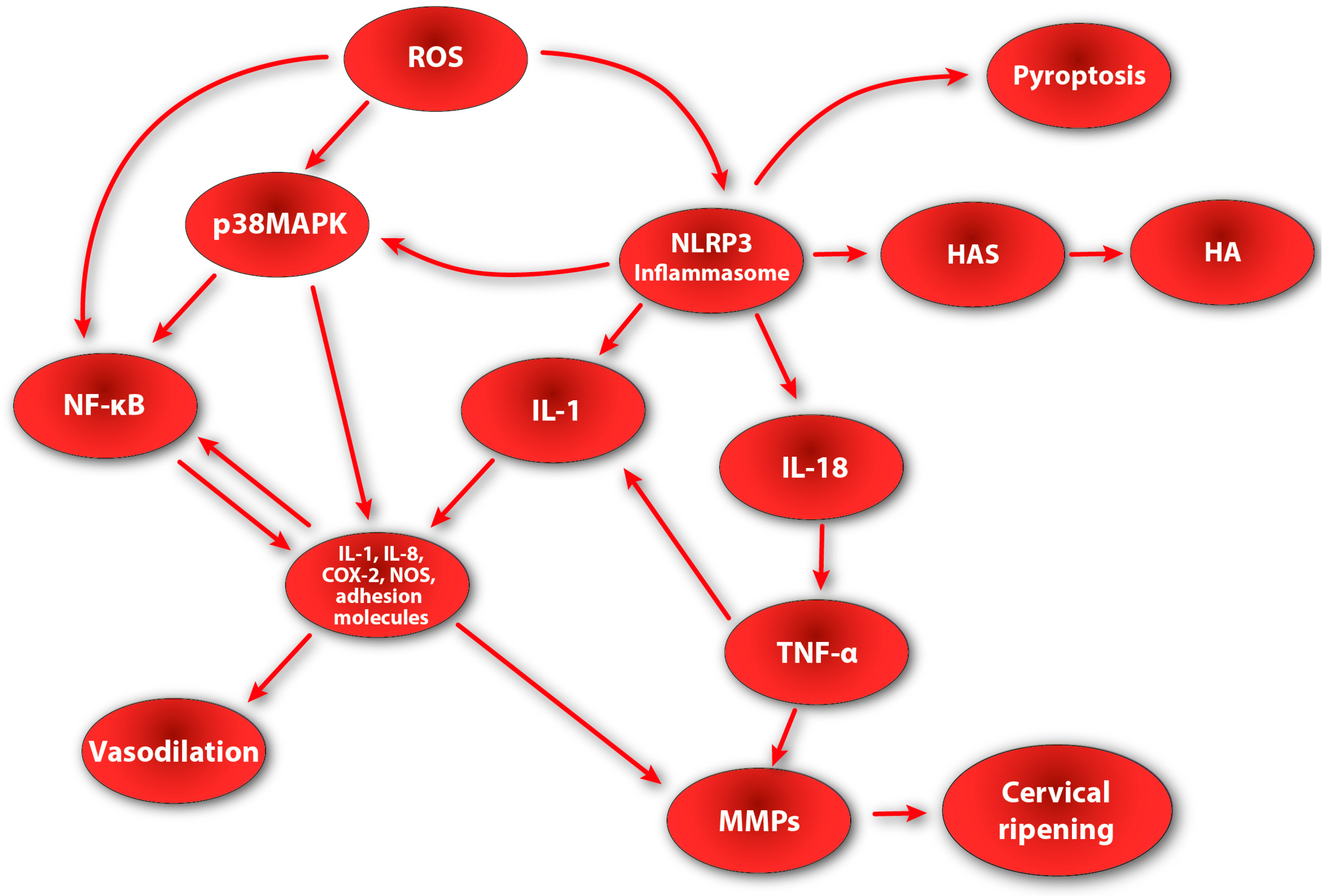
The presented data clearly indicate the active involvement of the inflammatory process in the ripening of the cervix. It appears that the assembly and activation of the NLRP3 inflammasome may also contribute to the inflammatory response during cervical remodeling. According to the research, in a term pregnancy, there is an increase in the concentration of inflammasome-dependent caspase-1 in maternal–fetal compartments such as fetal membranes, uterine muscle, and cervical tissue [154,155]. These data are consistent with reports showing that IL-1β (which is the main product of the inflammasome) is elevated in cervical tissue in women during the physiological process of cervical ripening and preterm labor [11,156]. Interestingly, in addition to the increased concentration of caspase-1, a significant increase in the expression of the adapter protein ASC and GSDMD was also noted in the cervical tissue [157,158,159,160,161]. The presence of the ASC adapter protein may indicate the occurrence of inflammasome assembly and activation processes in the cervical ECM during its ripening. Moreover, the expression of GSDMD may suggest that inflammatory changes in the cervical stroma may also be accompanied by the process of pyroptosis.
Recent studies shed additional light on the active involvement of the inflammasome in the molecular pathways of cervical ripening. To investigate the contribution of the NLRP3 inflammasome to the cervical ripening and onset of preterm labor, Nlrp3-sufficient and Nlrp3-deficient mice were used, which were administered LPS during late gestation. Strikingly, Nlrp3-sufficient mice displayed increased cervical distension as compared with Nlrp3-deficient mice. LPS induced cervical dilation in Nlrp3-deficient mice but to a much lesser extent, indicating impaired cervical distension [162]. Moreover, Nlrp3-sufficient mice showed increased expression patterns of genes associated with cervical structural remodeling, such as proteoglycans and HAS genes, suggesting that NLRP3 may directly participate in cervical stromal spatial reorganization [162,163]. Finally, the expression of key inflammatory cytokines such as IL-1β, IL-6, IL-8, and TNF was greatly up-regulated in Nlrp3-sufficient mice [162,163,164]. These data clearly seem to indicate the active and direct involvement of the active NLRP3 inflammasome in the metabolic pathways of cervical ripening.
We believe that the influence of the active NLRP3 inflammasome on cervical ripening is multilevel. Firstly, due to its influence on the expression of genes encoding enzymes involved in cervical remodeling (such as hyaluronidase synthase), it directly participates in structural changes in the cervical ECM, which translates into an increased dispersion of collagen and elastin fibers in the cervical stroma. Moreover, the active NLRP3 inflammasome has the ability to synthesize and secrete IL-1β and IL-18. By increasing the concentration of IL-1β, the secretion of other pro-inflammatory cytokines and MMPs increases, as well as the synthesis of PGs in the cervical stroma. In turn, IL-18 directly stimulates the activity of TNF—a cytokine that regulates other biochemical pathways of cervical ripening [165]. As a result of the action of TNF, the synthesis of other pro-inflammatory cytokines is additionally enhanced, and the nuclear translocation of NF-κB is additionally enhanced, which translates into an increase in the secretion and synthesis of NO, PGs, and cytokines. It is also worth mentioning the possible relationship between the NLRP3 inflammasome and p38MAPK. In the research, increased NLRP3 inflammasome activity (due to LPS activation) with subsequent elevation of IL-1β and caspase-1 was accompanied by greater activation of p38MAPK [166,167]. These data may suggest that NLRP3 and p38MAPK activation pathways may be interdependent and intertwined. Taking into account the fact that p38MAPK can be up-regulated by a variety of mediators (e.g., cytokines or ROS), we believe that due to the activity of NLRP3 and the subsequent increase in the secretion of cytokines (mainly IL-1β), p38MAPK is activated, which additionally translates into an increase in the local inflammatory reaction (Figure 2).
Taking all of the above into consideration, we believe that the NLRP3 inflammasome is actively involved both directly and indirectly in the molecular and biochemical pathways during cervical ripening. It is certain that its components and products are present in the stroma of the cervix during its ripening and are involved in both the inflammatory reaction and structural remodeling of the cervix. However, it does not appear that NLRP3 plays a primary role in initiating and conducting cervical ripening. The conducted research shows that the lack of NLRP3 does not inhibit cervical ripening, but only significantly impairs it [162]. On the other hand, the local inflammatory response appears to be one of the most important biochemical processes occurring both in preterm birth as well as in cervical ripening. Therefore, the presence of an active NLRP3 inflammasome seems to be crucial for the proper and uninterrupted course of these reactions. Summing up, we believe that the role of the NLRP3 inflammasome in the biochemical pathways of cervical ripening is to regulate, connect, and mutually drive several pathways that result in the development of a normal local inflammatory response leading to cervical architectural alteration. However, this interesting topic requires further, thorough research.
5. NLRP3 Inflammasome and Reactive Oxygen and Nitrogen Species in Cervical Tissue
Reactive oxygen species are free radicals that can be classified as reactive molecules that contain an unpaired, additional electron. The dominant forms are reactive nitrogen species (RNS) and reactive oxygen species (ROS) [168]. ROS are produced in a variety of biochemical pathways throughout the cell and organelles, such as the mitochondrial respiratory chain, peroxisomes, and endoplasmic reticulum (ER) [169]. Under physiological conditions, ROS are continuously secreted in small amounts in cellular compartments and are actively involved in maintaining homeostasis of the internal environment. ROS and RNS actively participate in intracellular signaling, redox regulations, activation of protein kinases, ion channel opening, and protein modifications [170,171,172]. Moreover, ROS are also antimicrobial factors that can directly destroy microbial pathogens [172]. The variance in the concentration of antioxidant and pro-oxidant molecules can promote the occurrence of oxidative stress [173]. The ratio of the reactive molecules can be altered by an increase in the concentration of pro-oxidants or by a decrease in antioxidant mechanisms. Regardless of the mechanism, in both mentioned situations, an accumulation of highly reactive molecules can be observed [174]. During excessive ROS synthesis, the antioxidant systems become insufficient, which impairs proper cell functioning. Redundancy of ROS (under pathological conditions) leads to catastrophic events such as DNA damage, lipid peroxidation, and cellular membrane disruption [173,174].
The dominant free radicals involved in cell homeostasis are the superoxide anion, hydroxyl ion, and peroxynitrite (ONOO−), with a dominance of the superoxide anion. Physiological sources of the superoxide anion include the mitochondrial respiratory chain, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, and cytochrome P450 [175,176,177]. Reactive nitrogen species (RNS) include nitric oxide (NO), nitrogen dioxide (NO2), and ONOO− [178]. NO is synthesized in the chemical reaction maintained by nitric oxide synthase (NOS), which requires NADPH as a cofactor and donor of additional electrons. NOS is an enzyme superfamily that consists of three isoforms: neuronal NOS (nNOS), inducible NOS (iNOS), and endothelial NOS (eNOS) [179]. Each of the isoforms is expressed in the cervical ECM, with a dominance of the inducible form [11,180,181]. Despite the fact that NO is a crucial molecule for proper cell homeostasis and cervical ripening, excessive secretion of NO can impair cell functionality [182].
The research clearly indicates the active involvement of reactive oxygen and nitrogen species in triggering and mediating the inflammatory response [183,184,185]. Moreover, it appears that ROS and RNS may also be actively involved in the biochemical and molecular pathways of cervical ripening. Referring to the research, ROS can directly stimulate the activity of p38MAPK in the cervical tissue, which is a strong mediator of the inflammatory reaction and also enhances the synthesis of other important factors involved in cervical ripening [145,186,187]. As mentioned earlier, stimulation of p38MAPK leads to increased expression of COX-2, pro-inflammatory cytokines, adhesion molecules, and MMPs [149]. Moreover, active p38MAPK has the ability to activate NF-κB, which leads to additional enhancement of the transcription of genes encoding factors such as NO, PGs, and pro-inflammatory cytokines [141]. In addition to activating NF-κB via p38MAPK (due to activation by ROS), reactive oxygen species can also directly stimulate the activation of NF-κB, which will result in intensified synthesis of the mentioned inflammatory reaction mediators. In its inactive form, NF-κB bonds with a specific NF-κB inhibitor, which can be expressed in three isoforms: IκBα, IκBβ, and IκBε [188]. NF-κB inhibitors are under strict regulation by IκB kinase (IKK). IKK can phosphorylate IκBα, subsequently leading to its degradation and allowing NF-κB to activate and translocate to the nucleus, where it becomes activated [188,189]. The excess ROS generated during the local inflammatory reaction can lead to the direct activation of protein kinase B (PKB) and NIK (NF-κB inducing kinase), whose role is to phosphorylate IKK. Once phosphorylated, IKK gains the ability to stimulate NF-κB. This leads to NF-κB activation, followed by an increase in the expression of factors regulating cervical ripening [190,191]. Moreover, pro-inflammatory cytokines involved in cervical remodeling can also stimulate ROS production. IL-1β and TNF (which are strong mediators of cervical ripening) can directly enhance the production of NADPH-oxidase-generated reactive oxygen species [192,193,194]. Subsequently, excess ROS may recruit other inflammatory mediators such as p38MAPK and NF-κB, which will lead to the boosting of inflammatory response in the cervical ECM. It appears that ROS and the NLRP3 inflammasome may also be associated with each other and act synergistically in the context of the development of a local inflammatory response. As mentioned earlier, both ROS and components of the NLRP3 inflammasome are present in cervical tissue during its remodeling period. Apart from the influence on NF-κB and p38MAPK, reactive oxygen species are potential inducers of NLRP3 inflammasome activation. Thioredoxin is believed to be involved in the activation of NLRP3 by ROS. Under resting conditions, thioredoxin-interacting protein (TXNIP) forms a complex with thioredoxin (TX) [195]. During excessive ROS production, an increased ROS concentration is sensed by the TXNIP-TX complex, leading to the dissociation of the complex. Then, liberated TXNIP can bind to the leucine-rich repeat of NLRP3, triggering the activation signal for NLPR3 inflammasome assembly [196,197].
Another possible mechanism for NLRP3 inflammasome activation by ROS is mitochondrial DNA (mtDNA) binding. Excessive production of ROS in mitochondria leads to the ROS binding to DNA strands, which leads to DNA damage and fragmentation. Then, free mtDNA fragments enter the cytoplasm, where they can directly initiate the assembly and activation of the NLRP3 inflammasome [196,198,199,200]. Activation of the NLRP3 inflammasome by ROS will result in increased secretion of caspase-1, IL-1β, and IL-18, which will translate into an increase in the inflammatory reaction and stimulation of further elements of its cascade, such as cytokines, PGs, MMPs, and NF-κB. Moreover, an active NLRP3 inflammasome can additionally enhance p38MAPK activity, which will further enhance the inflammatory response in cervical tissue. Furthermore, NLRP3-inflammasome-driven inflammation (induced by ROS) recruits inflammatory cells, including macrophages and neutrophils (whose concentration gradually increases in cervical tissue), which, in turn, leads to ROS production amplification, suggesting a feedback loop between ROS and the NLRP3 inflammasome. Taking the above into account, we believe that ROS not only directly participate in cervical ripening but are also responsible for the recruitment of additional, extremely important factors that regulate and stimulate the proper course of the local inflammatory reaction occurring in the cervical stroma. The presence of ROS seems to ensure the creation of a number of regulatory loops in which the components of the inflammatory process stimulate each other, leading to the intensification of changes occurring in the cervix.
6. Discussion
The main aim of this review was to provide the current state of knowledge of the possible involvement of the NLRP3 inflammasome in cervical ripening.
Cervical ripening is a derivative of the enzymatic breakdown of the extracellular matrix, mediated by a wide range of substances, which are strictly regulated by inflammatory and endocrine factors. There are numerous studies that describe the influence of specific factors on the cervical tissue in great detail. However, it has not yet been clearly postulated whether there is a single dominant factor or pathway that is responsible for triggering the complex biochemical pathways during cervical ripening. The data we described clearly indicate the active involvement of the NLRP3 inflammasome in the processes of cervical ripening. The NLRP3 inflammasome is intimately linked to almost every biochemical and molecular pathway involved in this complex process. After the activation and assembly of the NLRP3 inflammasome, IL-18 and IL-1β are secreted, which are potent mediators of cervical ripening. The secreted cytokines then activate a wide range of other mediators, such as MMP, NO, and PG. These substances, apart from directly affecting the remodeling of the cervix, may additionally intensify the inflammatory response. Starting from increasing vascular permeability and increasing the influx of inflammatory cells, through additional activation of NF-κB and p38MAPK, and ending with the activation of MMPs (decomposing collagen and elastin fibers), these substances lead to fundamental architectural changes in the cervix. Finally, substances whose synthesis is indirectly stimulated by the active NLRP3 inflammasome have the ability to additionally activate the NLRP3 inflammasome by acting as alarmins. Moreover, cytokines can enhance the production of ROS, which further amplify inflammasome activity. Taking the above into account, we believe that the NLRP3 inflammasome occupies a very important regulatory place in the processes of cervical ripening. Thanks to its activity, it is possible to connect and mutually stimulate molecular pathways dependent on cytokines, p38MAPK, NF-κB, and ROS.
In addition to its effects on the cervix, it appears that inflammasome activation may also influence uterine muscle contractility. As mentioned earlier, inflammasome components are present not only in the cervix but also in the myometrium during parturition. In previous work, the uterine tissue of Nlrp3-sufficient mice exhibited significantly increased expression of oxytocin receptors upon LPS injection as compared to the Nlrp3-deficient mice [163]. This may suggest that the assembly and activation of the NLRP3 inflammasome contributes to the increase in the expression of oxytocin receptors, which translates into increased contractility of the uterine myometrium during labor. When considering the function of the inflammasome, we should not forget the role of calcium ions in its activation. As mentioned earlier, increasing the concentration of calcium ions in the cell is one of the signals for the activation of the NLRP3 inflammasome. During labor, the contractility of the uterine myometrium depends largely on calcium ions. Both oxytocin and prostaglandins, by acting on their receptors, cause a significant increase in the concentration of calcium ions in the cytosol, which leads to the generation of muscle contraction [201,202]. We believe that an increase in the concentration of calcium ions in the cell (due to the interaction of PGs and oxytocin) additionally causes the activation of the NLRP3 inflammasome, which leads to an increase in the expression of oxytocin receptors and also increases the concentration of cytokines and caspase-1, which drives the development of a local inflammatory reaction. Moreover, in the research, the administration of MCC950, which is a specific inhibitor of the NLRP3 inflammasome, inhibited the occurrence of preterm birth and significantly reduced neonatal mortality [203,204]. Additionally, other studies conducted show that MCC950 could prove to be clinically safe and efficient for use in humans [205]. These data provide insights into the mechanisms of tissue dependencies in the context of preterm labor and indicate that targeting the NLRP3 pathway could prevent future adverse perinatal outcomes. Understanding how the NLRP3 inflammasome participates in molecular and biochemical pathways in terms of parturition and how to control excessive NLRP3 inflammasome activation is essential for the identification of new targets for the treatment of reproductive dysfunction. It seems that administering MCC950 to patients could inhibit the development of the inflammatory reaction occurring during cervical ripening in preterm labor. Taking into account the huge contribution of the inflammatory reaction to preterm birth, it seems that inhibiting one of its components, i.e., activation of the NLRP3 inflammasome, may contribute to inhibiting preterm birth, thus reducing neonatal complications. However, this interesting topic still requires more research to develop the mechanism-specific and safe treatments for pregnancy disorders.
7. Conclusions
Taking all of the above into consideration, we believe that the NLRP3 inflammasome holds a very important place in cervical biology. By participating in most biochemical pathways, it seems that the NLRP3 inflammasome may be responsible for mediating, regulating, and ensuring the proper functioning of all biochemical and molecular pathways that take place in the cervical tissue during its maturation. A comprehensive understanding of cervical biology seems to be crucial in the context of developing new therapeutic options to manage severe adverse perinatal complications such as preterm parturition. Despite our deep knowledge of the biochemical and molecular events occurring during the cervical ripening process, the hunt for the factors that trigger and modulate this process persists. An extensive understanding of the rules of cervical ripening seems to be pivotal in terms of labor induction. Greater understanding could provide us with resources to help women avoid dysfunctional parturition.
Author Contributions
Conceptualization, W.F. and M.W.S.; methodology, W.F. and M.W.S.; software, M.W.S.; validation, W.F.; formal analysis, M.W.S. and W.F.; investigation, W.F.; resources, M.W.S.; data curation, W.F. and M.W.S.; writing—original draft preparation, W.F.; writing—review and editing, W.F. and M.W.S.; visualization, M.W.S.; supervision, W.F. and M.W.S.; project administration, W.F. and M.W.S.; funding acquisition, M.W.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Martyn, F.; McAuliffe, F.M.; Wingfield, M. The role of the cervix in fertility: Is it time for a reappraisal? Hum. Reprod. 2014, 29, 2092–2098. [Google Scholar] [CrossRef] [PubMed]
- Sennström, M.B.; Ekman, G.; Westergren-Thorsson, G.; Malmström, A.; Byström, B.; Endrésen, U.; Mlambo, N.; Norman, M.; Ståbi, B. Annelie Brauner, Human cervical ripening, an inflammatory process mediated by cytokines. Mol. Hum. Reprod. 2000, 6, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Ekman-Ordeberg, G.; Stjernholm, Y.; Wang, H.; Stygar, D.; Sahlin, L. Endocrine regulation of cervical ripening in humans—Potential roles for gonadal steroids and insulin-like growth factor-I. Steroids 2003, 68, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Gross, O.; Thomas, C.J.; Guarda, G.; Tschopp, J. The inflammasome: An integrated view. Immunol. Rev. 2011, 243, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Plazyo, O.; Unkel, R.; Leng, Y.; Than, N.G.; Chaiworapongsa, T.; Panaitescu, B.; Dong, Z.; et al. A Role for the Inflammasome in Spontaneous Preterm Labor with Acute Histologic Chorioamnionitis. Reprod. Sci. 2017, 24, 1382–1401. [Google Scholar] [CrossRef]
- Spencer, T.E.; Hayashi, K.; Hu, J.; Carpenter, K.D. Comparative developmental biology of the mammalian uterus. Curr. Top. Dev. Biol. 2005, 68, 85–122. [Google Scholar] [CrossRef] [PubMed]
- Myers, K.M.; Feltovich, H.; Mazza, E.; Vink, J.; Bajka, M.; Wapner, R.J.; Hall, T.J.; House, M. The mechanical role of the cervix in pregnancy. J. Biomech. 2015, 48, 1511–1523. [Google Scholar] [CrossRef]
- Leppert, P.C. Anatomy and physiology of cervical ripening. Clin. Obstet. Gynecol. 1995, 38, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Vink, J.Y.; Qin, S.; Brock, C.O.; Zork, N.M.; Feltovich, H.M.; Chen, X.; Urie, P.; Myers, K.M.; Hall, T.J.; Wapner, R.; et al. A new paradigm for the role of smooth muscle cells in the human cervix. Am. J. Obstet. Gynecol. 2016, 215, 478.e1–478.e11. [Google Scholar] [CrossRef]
- Nicoll, A. The Physiology of Cervical Ripening and the Induction of Labour: A Potential Role for the Nitric Oxide Donor Isosorbide Mononitrate; University of Glasgow: Glasgow, UK, 2001. [Google Scholar]
- Iwahashi, M.; Muragaki, Y.; Ooshima, A.; Umesaki, N. Decreased type I collagen expression in human uterine cervix during pregnancy. J. Clin. Endocrinol. Metab. 2003, 88, 2231–2235. [Google Scholar] [CrossRef]
- Wang, Q.; Chi, L. The Alterations and Roles of Glycosaminoglycans in Human Diseases. Polymers 2022, 14, 5014. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Yao, W.; Harris, W.B.R.; Vink, J.Y.; Myers, K.M.; Donnelly, E. Characterization of the collagen microstructural organization of human cervical tissue. Reproduction 2018, 156, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Oxlund, B.S.; Ørtoft, G.; Brüel, A.; Danielsen, C.C.; Bor, P.; Oxlund, H.; Uldbjerg, N. Collagen concentration and biomechanical properties of samples from the lower uterine cervix in relation to age and parity in non-pregnant women. Reprod. Biol. Endocrinol. 2010, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Halper, J. Proteoglycans and diseases of soft tissues. Adv. Exp. Med. Biol. 2014, 802, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Akgul, Y.; Holt, R.; Mummert, M.; Word, A.; Mahendroo, M. Dynamic changes in cervical glycosaminoglycan composition during normal pregnancy and preterm birth. Endocrinology 2012, 153, 3493–3503. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Jayyosi, C.; Lee, N.; Mahendroo, M.; Myers, K.M. Mechanics of cervical remodelling: Insights from rodent models of pregnancy. Interface Focus 2019, 9, 20190026. [Google Scholar] [CrossRef] [PubMed]
- Ruscheinsky, M.; De la Motte, C.; Mahendroo, M. Hyaluronan and its binding proteins during cervical ripening and parturition: Dynamic changes in size, distribution and temporal sequence. Matrix Biol. 2008, 27, 487–497. [Google Scholar] [CrossRef]
- Schmelzer, C.E.H.; Duca, L. Elastic fibers: Formation, function, and fate during aging and disease. FEBS J. 2022, 289, 3704–3730. [Google Scholar] [CrossRef] [PubMed]
- Nallasamy, S.; Yoshida, K.; Akins, M.; Myers, K.; Iozzo, R.; Mahendroo, M. Steroid Hormones Are Key Modulators of Tissue Mechanical Function via Regulation of Collagen and Elastic Fibers. Endocrinology 2017, 158, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Leppert, P.C.; Keller, S.; Cerreta, J.; Hosannah, Y.; Mandl, I. The content of elastin in the uterine cervix. Arch. Biochem. Biophys. 1983, 222, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Levine, L.D. Cervical ripening: Why we do what we do. Semin. Perinatol. 2020, 44, 151216. [Google Scholar] [CrossRef]
- Wheeler, V.; Hoffman, A.; Bybel, M. Cervical Ripening and Induction of Labor. Am. Fam. Physician 2022, 105, 177–186, Erratum in Am. Fam. Physician 2022, 106, 121. [Google Scholar] [PubMed]
- El Maradny, E.; Kanayama, N.; Kobayashi, H.; Hossain, B.; Khatun, S.; Liping, S.; Kobayashi, T.; Terao, T. The role of hyaluronic acid as a mediator and regulator of cervical ripening. Hum. Reprod. 1997, 12, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Uldbjerg, N.; Ekman, G.; Malmström, A.; Olsson, K.; Ulmsten, U. Ripening of the human uterine cervix related to changes in collagen, glycosaminoglycans, and collagenolytic activity. Am. J. Obstet. Gynecol. 1983, 147, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Di, X.; Wang, L.; Zhang, W.; Feng, Y.; Shi, S.Q.; Garfield, R.E.; Liu, H. Evaluation of cervical maturity by cervical collagen measurement using light-induced fluorescence (LIF) during pregnancy. J. Int. Med. Res. 2020, 48, 300060520964006. [Google Scholar] [CrossRef] [PubMed]
- Maul, H.; Mackay, L.; Garfield, R.E. Cervical ripening: Biochemical, molecular, and clinical considerations. Clin. Obstet. Gynecol. 2006, 49, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuña, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Sennström, M.B.; Brauner, A.; Byström, B.; Malmström, A.; Ekman, G. Matrix metalloproteinase-8 correlates with the cervical ripening process in humans. Acta Obstet. Gynecol. Scand. 2003, 82, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Stygar, D.; Wang, H.; Vladic, Y.S.; Ekman, G.; Eriksson, H.; Sahlin, L. Increased level of matrix metalloproteinases 2 and 9 in the ripening process of the human cervix. Biol. Reprod. 2002, 67, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Jung, K.L.; Oh, S.Y.; Kim, J.H.; Roh, C.R. Cervicovaginal matrix metalloproteinase-9 and cervical ripening in human term parturition. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 142, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.Y.; Mao, Q.X.; Luo, J.H. Dynamic expression of matrix metalloproteinases 2 and 8 in rat cervix. Nan Fang Yi Ke Da Xue Xue Bao 2009, 29, 2205–2207. (In Chinese) [Google Scholar] [PubMed]
- Bollapragada, S.; Youssef, R.; Jordan, F.; Greer, I.; Norman, J.; Nelson, S. Term labor is associated with a core inflammatory response in human fetal membranes, myometrium, and cervix. Am. J. Obstet. Gynecol. 2009, 200, 104.e1–104.e11, Erratum in Am. J. Obstet. Gynecol. 2009, 201, 214. Bollopragada, Shrikant [corrected to Bollapragada, Shrikant]. [Google Scholar] [CrossRef] [PubMed]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Fingleton, B. Matrix metalloproteinases as regulators of inflammatory processes. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2036–2042. [Google Scholar] [CrossRef] [PubMed]
- Barnett, K.C.; Li, S.; Liang, K.; Ting, J.P. A 360° view of the inflammasome: Mechanisms of activation, cell death, and diseases. Cell 2023, 186, 2288–2312. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.G.; Dash, P.; Aldridge, J.R., Jr.; Ellebedy, A.H.; Reynolds, C.; Funk, A.J.; Martin, W.J.; Lamkanfi, M.; Webby, R.J.; Boyd, K.L.; et al. The intracellular sensor NLRP3 mediates key innate and healing responses to influenza A virus via the regulation of caspase-1. Immunity 2009, 30, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Allen, I.C.; Scull, M.A.; Moore, C.B.; Holl, E.K.; McElvania-TeKippe, E.; Taxman, D.J.; Guthrie, E.H.; Pickles, R.J.; Ting, J.P. The NLRP3 inflammasome mediates in vivo innate immunity to influenza A virus through recognition of viral RNA. Immunity 2009, 30, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Gross, O.; Poeck, H.; Bscheider, M.; Dostert, C.; Hannesschläger, N.; Endres, S.; Hartmann, G.; Tardivel, A.; Schweighoffer, E.; Tybulewicz, V.; et al. Syk kinase signalling couples to the Nlrp3 inflammasome for anti-fungal host defence. Nature 2009, 459, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef]
- Oroz, J.; Barrera-Vilarmau, S.; Alfonso, C.; Rivas, G.; de Alba, E. ASC Pyrin Domain Self-associates and Binds NLRP3 Protein Using Equivalent Binding Interfaces. J. Biol. Chem. 2016, 291, 19487–19501. [Google Scholar] [CrossRef] [PubMed]
- Manji, G.A.; Wang, L.; Geddes, B.J.; Brown, M.; Merriam, S.; Al-Garawi, A.; Mak, S.; Lora, J.M.; Briskin, M.; Jurman, M.; et al. PYPAF1, a PYRIN-containing Apaf1-like protein that assembles with ASC and regulates activation of NF-kappa B. J. Biol. Chem. 2002, 277, 11570–11575. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Immunological and inflammatory functions of the interleukin-1 family. Annu. Rev. Immunol. 2009, 27, 519–550. [Google Scholar] [CrossRef] [PubMed]
- Fink, S.L.; Cookson, B.T. Caspase-1-dependent pore formation during pyroptosis leads to osmotic lysis of infected host macrophages. Cell. Microbiol. 2006, 8, 1812–1825. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Miao, E.A.; Leaf, I.A.; Treuting, P.M.; Mao, D.P.; Dors, M.; Sarkar, A.; Warren, S.E.; Wewers, M.D.; Aderem, A. Caspase-1-induced pyroptosis is an innate immune effector mechanism against intracellular bacteria. Nat. Immunol. 2010, 11, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Menu, P.; Vince, J.E. The NLRP3 inflammasome in health and disease: The good, the bad and the ugly. Clin. Exp. Immunol. 2011, 166, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Kanneganti, T.D. Nlrp3: An immune sensor of cellular stress and infection. Int. J. Biochem. Cell Biol. 2010, 42, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Warner, N.; Viani, K.; Nuñez, G. Function of Nod-like receptors in microbial recognition and host defense. Immunol. Rev. 2009, 227, 106–128. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.A.; Bergstralh, D.T.; Wang, Y.; Willingham, S.B.; Ye, Z.; Zimmermann, A.G.; Ting, J.P. Cryopyrin/NALP3 binds ATP/dATP, is an ATPase, and requires ATP binding to mediate inflammatory signaling. Proc. Natl. Acad. Sci. USA 2007, 104, 8041–8046. [Google Scholar] [CrossRef] [PubMed]
- Proell, M.; Gerlic, M.; Mace, P.D.; Reed, J.C.; Riedl, S.J. The CARD plays a critical role in ASC foci formation and inflammasome signalling. Biochem. J. 2013, 449, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Elizagaray, M.L.; Gomes, M.T.R.; Guimaraes, E.S.; Rumbo, M.; Hozbor, D.F.; Oliveira, S.C.; Moreno, G. Canonical and Non-canonical Inflammasome Activation by Outer Membrane Vesicles Derived from Bordetella pertussis. Front. Immunol. 2020, 11, 1879. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Hara, H.; Núñez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Gurung, P.; Anand, P.K.; Malireddi, R.K.; Vande Walle, L.; Van Opdenbosch, N.; Dillon, C.P.; Weinlich, R.; Green, D.R.; Lamkanfi, M.; Kanneganti, T.D. FADD and caspase-8 mediate priming and activation of the canonical and noncanonical Nlrp3 inflammasomes. J. Immunol. 2014, 192, 1835–1846. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Li, Q.; Xu, G.; Xiao, X.; Bai, Z. The mechanism of NLRP3 inflammasome activation and its pharmacological inhibitors. Front. Immunol. 2023, 13, 1109938. [Google Scholar] [CrossRef]
- Fang, X.; Wang, Y.; Zhang, Y.; Li, Y.; Kwak-Kim, J.; Wu, L. NLRP3 Inflammasome and Its Critical Role in Gynecological Disorders and Obstetrical Complications. Front. Immunol. 2021, 11, 555826. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Ismail, N. Non-Canonical Inflammasome Pathway: The Role of Cell Death and Inflammation in Ehrlichiosis. Cells 2023, 12, 2597. [Google Scholar] [CrossRef] [PubMed]
- Santa Cruz Garcia, A.B.; Schnur, K.P.; Malik, A.B.; Mo, G.C.H. Gasdermin D pores are dynamically regulated by local phosphoinositide circuitry. Nat. Commun. 2022, 13, 52. [Google Scholar] [CrossRef] [PubMed]
- Devant, P.; Kagan, J.C. Molecular mechanisms of gasdermin D pore-forming activity. Nat. Immunol. 2023, 24, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.S.; Sohn, D.H. Damage-Associated Molecular Patterns in Inflammatory Diseases. Immune Netw. 2018, 18, e27. [Google Scholar] [CrossRef] [PubMed]
- Eigenbrod, T.; Dalpke, A.H. Bacterial RNA: An Underestimated Stimulus for Innate Immune Responses. J. Immunol. 2015, 195, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Ghosh, S.; Monks, B.; DeOliveira, R.B.; Tzeng, T.C.; Kalantari, P.; Nandy, A.; Bhattacharjee, B.; Chan, J.; Ferreira, F.; et al. RNA and β-hemolysin of group B Streptococcus induce interleukin-1β (IL-1β) by activating NLRP3 inflammasomes in mouse macrophages. J. Biol. Chem. 2014, 289, 13701–13705. [Google Scholar] [CrossRef] [PubMed]
- Sha, W.; Mitoma, H.; Hanabuchi, S.; Bao, M.; Weng, L.; Sugimoto, N.; Liu, Y.; Zhang, Z.; Zhong, J.; Sun, B.; et al. Human NLRP3 inflammasome senses multiple types of bacterial RNAs. Proc. Natl. Acad. Sci. USA 2014, 111, 16059–16064. [Google Scholar] [CrossRef] [PubMed]
- Volonté, C.; Apolloni, S.; Skaper, S.D.; Burnstock, G. P2X7 receptors: Channels, pores and more. CNS Neurol. Disord. Drug Targets 2012, 11, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Eigenbrod, T.; Muñoz-Planillo, R.; Ozkurede, U.; Kim, Y.G.; Arindam, C.; Gale MJr Silverman, R.H.; Colonna, M.; Akira, S.; Núñez, G. Cytosolic double-stranded RNA activates the NLRP3 inflammasome via MAVS-induced membrane permeabilization and K+ efflux. J. Immunol. 2014, 193, 4214–4222. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zeng, M.Y.; Yang, D.; Motro, B.; Núñez, G. NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux. Nature 2016, 530, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Burgk, J.L.; Chauhan, D.; Schmidt, T.; Ebert, T.S.; Reinhardt, J.; Endl, E.; Hornung, V. A Genome-wide CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) Screen Identifies NEK7 as an Essential Component of NLRP3 Inflammasome Activation. J. Biol. Chem. 2016, 291, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wang, Y.; Li, X.; Zhan, X.; Tang, M.; Fina, M.; Su, L.; Pratt, D.; Bu, C.H.; Hildebrand, S.; et al. NLRP3 activation and mitosis are mutually exclusive events coordinated by NEK7, a new inflammasome component. Nat. Immunol. 2016, 17, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lu, L.; Li, L. NEK7: A novel promising therapy target for NLRP3-related inflammatory diseases. Acta Biochim. Biophys. Sin. 2016, 48, 966–968. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Lang, X.; Xu, C.; Wang, X.; Gong, T.; Yang, Y.; Cui, J.; Bai, L.; Wang, J.; Jiang, W.; et al. CLICs-dependent chloride efflux is an essential and proximal upstream event for NLRP3 inflammasome activation. Nat. Commun. 2017, 8, 202. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Yang, Y.; Jin, T.; Jiang, W.; Zhou, R. Orchestration of NLRP3 Inflammasome Activation by Ion Fluxes. Trends Immunol. 2018, 39, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.S.; Subramanian, N.; Kim, A.I.; Aksentijevich, I.; Goldbach-Mansky, R.; Sacks, D.B.; Germain, R.N.; Kastner, D.L.; Chae, J.J. The calcium-sensing receptor regulates the NLRP3 inflammasome through Ca2+ and cAMP. Nature 2012, 492, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Rossol, M.; Pierer, M.; Raulien, N.; Quandt, D.; Meusch, U.; Rothe, K.; Schubert, K.; Schöneberg, T.; Schaefer, M.; Krügel, U.; et al. Extracellular Ca2+ is a danger signal activating the NLRP3 inflammasome through G protein-coupled calcium sensing receptors. Nat. Commun. 2012, 3, 1329. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, C.H.; Carroll, R.G.; Wei, P.; Jones, K.A.; Rice, A.P. Tat-associated kinase, TAK, activity is regulated by distinct mechanisms in peripheral blood lymphocytes and promonocytic cell lines. J. Virol. 1998, 72, 9881–9888. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, K.; Saharinen, P.; Pesu, M.; Holt, V.E., 3rd; Silvennoinen, O.; O’Shea, J.J. The Janus kinases (Jaks). Genome Biol. 2004, 5, 253. [Google Scholar] [CrossRef]
- Elliott, E.I.; Sutterwala, F.S. Initiation and perpetuation of NLRP3 inflammasome activation and assembly. Immunol. Rev. 2015, 265, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Verhoef, P.A.; Kertesy, S.B.; Lundberg, K.; Kahlenberg, J.M.; Dubyak, G.R. Inhibitory effects of chloride on the activation of caspase-1, IL-1beta secretion, and cytolysis by the P2X7 receptor. J. Immunol. 2005, 175, 7623–7634. [Google Scholar] [CrossRef] [PubMed]
- Perregaux, D.G.; Laliberte, R.E.; Gabel, C.A. Human monocyte interleukin-1beta posttranslational processing. Evidence of a volume-regulated response. J. Biol. Chem. 1996, 271, 29830–29838. [Google Scholar] [CrossRef] [PubMed]
- Compan, V.; Baroja-Mazo, A.; López-Castejón, G.; Gomez, A.I.; Martínez, C.M.; Angosto, D.; Montero, M.T.; Herranz, A.S.; Bazán, E.; Reimers, D.; et al. Cell volume regulation modulates NLRP3 inflammasome activation. Immunity. 2012, 37, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Domingo-Fernández, R.; Coll, R.C.; Kearney, J.; Breit, S.; O’Neill, L.A.J. The intracellular chloride channel proteins CLIC1 and CLIC4 induce IL-1β transcription and activate the NLRP3 inflammasome. J. Biol. Chem. 2017, 292, 12077–12087. [Google Scholar] [CrossRef] [PubMed]
- Green, J.P.; Yu, S.; Martín-Sánchez, F.; Pelegrin, P.; Lopez-Castejon, G.; Lawrence, C.B.; Brough, D. Chloride regulates dynamic NLRP3-dependent ASC oligomerization and inflammasome priming. Proc. Natl. Acad. Sci. USA 2018, 115, E9371–E9380. [Google Scholar] [CrossRef] [PubMed]
- Hagar, J.A.; Powell, D.A.; Aachoui, Y.; Ernst, R.K.; Miao, E.A. Cytoplasmic LPS activates caspase-11: Implications in TLR4-independent endotoxic shock. Science 2013, 341, 1250–1253. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Wong, M.T.; Stowe, I.B.; Ramani, S.R.; Gonzalez, L.C.; Akashi-Takamura, S.; Miyake, K.; Zhang, J.; Lee, W.P.; Muszyński, A.; et al. Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science 2013, 341, 1246–1249. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Warming, S.; Lamkanfi, M.; Vande Walle, L.; Louie, S.; Dong, J.; Newton, K.; Qu, Y.; Liu, J.; Heldens, S.; et al. Non-canonical inflammasome activation targets caspase-11. Nature 2011, 479, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.J.; Boucher, D.; Bierschenk, D.; Tebartz, C.; Whitney, P.G.; D’Silva, D.B.; Tanzer, M.C.; Monteleone, M.; Robertson, A.A.; Cooper, M.A.; et al. NLRP3 inflammasome activation downstream of cytoplasmic LPS recognition by both caspase-4 and caspase-5. Eur. J. Immunol. 2015, 45, 2918–2926. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Miura, M.; Jung, Y.K.; Zhu, H.; Li, E.; Yuan, J. Murine caspase-11, an ICE-interacting protease, is essential for the activation of ICE. Cell 1998, 92, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Rühl, S.; Broz, P. Caspase-11 activates a canonical NLRP3 inflammasome by promoting K+ efflux. Eur. J. Immunol. 2015, 45, 2927–2936. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, I.; Tan, Y.; Di Gioia, M.; Broggi, A.; Ruan, J.; Shi, J.; Donado, C.A.; Shao, F.; Wu, H.; Springstead, J.R.; et al. An endogenous caspase-11 ligand elicits interleukin-1 release from living dendritic cells. Science 2016, 352, 1232–1236. [Google Scholar] [CrossRef] [PubMed]
- He, W.T.; Wan, H.; Hu, L.; Chen, P.; Wang, X.; Huang, Z.; Yang, Z.H.; Zhong, C.Q.; Han, J. Gasdermin D is an executor of pyroptosis and required for interleukin-1β secretion. Cell Res. 2015, 25, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.; Sun, H.; Wang, D.C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lopez, N.; Guilbert, L.J.; Olson, D.M. Invasion of the leukocytes into the fetal-maternal interface during pregnancy. J. Leukoc. Biol. 2010, 88, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Stjernholm-Vladic, Y.; Stygar, D.; Mansson, C.; Masironi, B.; Akerberg, S.; Wang, H.; Ekman-Ordeberg, G.; Sahlin, L. Factors involved in the inflammatory events of cervical ripening in humans. Reprod. Biol. Endocrinol. 2004, 2, 74. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.W. Inflammatory mediators and cervical ripening. J. Reprod. Immunol. 2002, 57, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Shuai, H.; Luo, X.; Wang, X.; Guan, B. The clinical and prognostic value of CXCL8 in cervical carcinoma patients: Immunohistochemical analysis. Biosci. Rep. 2017, 37, BSR20171021. [Google Scholar] [CrossRef] [PubMed]
- Rhee, K.Y.; Goetzl, L.; Unal, R.; Cierny, J.; Flood, P. The Relationship between Plasma Inflammatory Cytokines and Labor Pain. Anesth. Analg. 2015, 121, 748–751. [Google Scholar] [CrossRef] [PubMed]
- el Maradny, E.; Kanayama, N.; Halim, A.; Maehara, K.; Sumimoto, K.; Terao, T. The effect of interleukin-1 in rabbit cervical ripening. Eur. J. Obstet. Gynecol. Reprod. Biol. 1995, 60, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Watari, M.; Watari, H.; DiSanto, M.E.; Chacko, S.; Shi, G.P.; Strauss, J.F., 3rd. Pro-inflammatory cytokines induce expression of matrix-metabolizing enzymes in human cervical smooth muscle cells. Am. J. Pathol. 1999, 154, 1755–1762. [Google Scholar] [CrossRef]
- Boraschi, D. What Is IL-1 for? The Functions of Interleukin-1 Across Evolution. Front. Immunol. 2022, 13, 872155. [Google Scholar] [CrossRef]
- Ristimäki, A.; Garfinkel, S.; Wessendorf, J.; Maciag, T.; Hla, T. Induction of cyclooxygenase-2 by interleukin-1 alpha. Evidence for post-transcriptional regulation. J. Biol. Chem. 1994, 269, 11769–11775. [Google Scholar] [CrossRef] [PubMed]
- Ogata, S.; Kubota, Y.; Yamashiro, T.; Takeuchi, H.; Ninomiya, T.; Suyama, Y.; Shirasuna, K. Signaling pathways regulating IL-1alpha-induced COX-2 expression. J. Dent. Res. 2007, 86, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Mertin, J.; Stackpoole, A.; Shumway, S.J. Prostaglandins and cell-mediated immunity. The role of prostaglandin E1 in the induction of host-versus-graft and graft-versus-host reactions in mice. Transplantation 1984, 37, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; DeWitt, D.L.; McNeely, T.B.; Wahl, S.M.; Wahl, L.M. Secretory leukocyte protease inhibitor suppresses the production of monocyte prostaglandin H synthase-2, prostaglandin E2, and matrix metalloproteinases. J. Clin. Investig. 1997, 99, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, J.D.; Kashiwagi, K.; Boyle, D.; Kashiwagi, F.; Firestein, G.S.; Weinreb, R.N. Prostaglandins increase proMMP-1 and proMMP-3 secretion by human ciliary smooth muscle cells. Curr. Eye Res. 1996, 15, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Karli, S.; Ayala-Haedo, J.A.; Feuer, W.J.; Fernandez, M.; Dubovy, S.; Wester, S.T. Effect of prostaglandin analogs on matrix metalloproteinases and tissue inhibitor of metalloproteinases in eyelid muscle specimens. Clin. Ophthalmol. 2018, 12, 2039–2046. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, I.; Zaragoza, D.B.; Guilbert, L.; Robertson, S.A.; Mitchell, B.F.; Olson, D.M. Inflammatory processes in preterm and term parturition. J. Reprod. Immunol. 2008, 79, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Ehsani, V.; Mortazavi, M.; Ghorban, K.; Dadmanesh, M.; Bahramabadi, R.; Rezayati, M.T.; Javadi-Moghadam, E.; Rezaei, Z.; Sabzali, Z.; Fatemi, I.; et al. Role of maternal interleukin-8 (IL-8) in normal-term birth in the human. Reprod. Fertil. Dev. 2019, 31, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Osmers, R.G.; Bläser, J.; Kuhn, W.; Tschesche, H. Interleukin-8 synthesis and the onset of labor. Obstet. Gynecol. 1995, 86, 223–229. [Google Scholar] [CrossRef]
- Winkler, M.; Fischer, D.C.; Hlubek, M.; van de Leur, E.; Haubeck, H.D.; Rath, W. Interleukin-1beta and interleukin-8 concentrations in the lower uterine segment during parturition at term. Obstet. Gynecol. 1998, 91, 945–949. [Google Scholar] [CrossRef]
- Khatun, S.; Kanayama, N.; Md Belayet, H.; Yonezawa, M.; Kobayashi, T.; Terao, T. Interleukin-8 potentiates the effect of interleukin-1-induced uterine contractions. Hum. Reprod. 1999, 14, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Lattey, K.R.; Kelly, A.J. Nitric oxide donors for cervical ripening and induction of labour. Cochrane Database Syst. Rev. 2016, 12, CD006901. [Google Scholar] [CrossRef] [PubMed]
- Tschugguel, W.; Schneeberger, C.; Lass, H.; Stonek, F.; Zaghlula, M.B.; Czerwenka, K.; Schatten, C.; Kaider, A.; Husslein, P.; Huber, J.C. Human cervical ripening is associated with an increase in cervical inducible nitric oxide synthase expression. Biol. Reprod. 1999, 60, 1367–1372. [Google Scholar] [CrossRef]
- Laskin, D.L.; Pendino, K.J. Macrophages and inflammatory mediators in tissue injury. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 655–677. [Google Scholar] [CrossRef] [PubMed]
- Chwalisz, K.; Shao-Qing, S.; Garfield, R.E.; Beier, H.M. Cervical ripening in guinea-pigs after a local application of nitric oxide. Hum. Reprod. 1997, 12, 2093–2101. [Google Scholar] [CrossRef] [PubMed]
- Tiboni, G.M.; Giampietro, F. Inhibition of nitric oxide synthesis causes preterm delivery in the mouse. Hum. Reprod. 2000, 15, 1838–1842. [Google Scholar] [CrossRef] [PubMed]
- Väisänen-Tommiska, M.R. Nitric oxide in the human uterine cervix: Endogenous ripening factor. Ann. Med. 2008, 40, 45–55. [Google Scholar] [CrossRef]
- Sasaki, K.; Hattori, T.; Fujisawa, T.; Takahashi, K.; Inoue, H.; Takigawa, M. Nitric oxide mediates interleukin-1-induced gene expression of matrix metalloproteinases and basic fibroblast growth factor in cultured rabbit articular chondrocytes. J. Biochem. 1998, 123, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Corriveau, C.C.; Madara, P.J.; Van Dervort, A.L.; Tropea, M.M.; Wesley, R.A.; Danner, R.L. Effects of nitric oxide on chemotaxis and endotoxin-induced interleukin-8 production in human neutrophils. J. Infect. Dis. 1998, 177, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ohigashi, H. Targeting NOX, INOS and COX-2 in inflammatory cells: Chemoprevention using food phytochemicals. Int. J. Cancer. 2007, 121, 2357–2363. [Google Scholar] [CrossRef] [PubMed]
- Ledingham, M.A.; Denison, F.C.; Kelly, R.W.; Young, A.; Norman, J.E. Nitric oxide donors stimulate prostaglandin F(2alpha) and inhibit thromboxane B(2) production in the human cervix during the first trimester of pregnancy. Mol. Hum. Reprod. 1999, 5, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A. Nitric Oxide Synthase and Cyclooxygenase Pathways: A Complex Interplay in Cellular Signaling. Curr. Med. Chem. 2016, 23, 2559–2578. [Google Scholar] [CrossRef] [PubMed]
- Jang, D.I.; Lee, A.H.; Shin, H.Y.; Song, H.R.; Park, J.H.; Kang, T.B.; Lee, S.R.; Yang, S.H. The Role of Tumor Necrosis Factor Alpha (TNF-α) in Autoimmune Disease and Current TNF-α Inhibitors in Therapeutics. Int. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, E.; Filipovich, Y.; Mahendroo, M. Signaling via the type I IL-1 and TNF receptors is necessary for bacterially induced preterm labor in a murine model. Am. J. Obstet. Gynecol. 2006, 194, 1334–1340. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, J.; Lara-Reyna, S.; Jarosz-Griffiths, H.; McDermott, M. Tumour necrosis factor signalling in health and disease. F1000Research 2019, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Arias-Salvatierra, D.; Silbergeld, E.K.; Acosta-Saavedra, L.C.; Calderon-Aranda, E.S. Role of nitric oxide produced by iNOS through NF-κB pathway in migration of cerebellar granule neurons induced by Lipopolysaccharide. Cell Signal. 2011, 23, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.W.; Kim, H.; Kim, K.H. Nuclear factor-kappaB regulates cyclooxygenase-2 expression and cell proliferation in human gastric cancer cells. Lab. Investig. 2001, 81, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-kappaB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol. 2006, 72, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Allport, V.C.; Pieber, D.; Slater, D.M.; Newton, R.; White, J.O.; Bennett, P.R. Human labour is associated with nuclear factor-kappaB activity which mediates cyclo-oxygenase-2 expression and is involved with the ‘functional progesterone withdrawal’. Mol. Hum. Reprod. 2001, 7, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Madrid, L.V.; Wang, C.Y.; Guttridge, D.C.; Schottelius, A.J.; Baldwin, A.S., Jr.; Mayo, M.W. Akt suppresses apoptosis by stimulating the transactivation potential of the RelA/p65 subunit of NF-kappaB. Mol. Cell. Biol. 2000, 20, 1626–1638. [Google Scholar] [CrossRef] [PubMed]
- Clancy, R.M.; Gomez, P.F.; Abramson, S.B. Nitric oxide sustains nuclear factor kappaB activation in cytokine-stimulated chondrocytes. Osteoarthr. Cartil. 2004, 12, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kim, S.C.; Yu, T.; Yi, Y.S.; Rhee, M.H.; Sung, G.H.; Yoo, B.C.; Cho, J.Y. Functional roles of p38 mitogen-activated protein kinase in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2014, 2014, 352371. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.B.; Cameron, P.M.; O’Keefe, S.J.; Frantz-Wattley, B.; Thompson, J.; O’Neill, E.A.; Tennis, T.; Liu, L.; Becker, J.W.; Scapin, G. The three-dimensional structure of MAP kinase p38beta: Different features of the ATP-binding site in p38beta compared with p38alpha. Acta Crystallogr. Sect. D Struct. Biol. 2009, 65, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Stjernholm, Y.V. Plasma membrane receptor mediated MAPK signaling pathways are activated in human uterine cervix at parturition. Reprod. Biol. Endocrinol. 2007, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Cindrova-Davies, T.; Spasic-Boskovic, O.; Jauniaux, E.; Charnock-Jones, D.S.; Burton, G.J. Nuclear factor-kappa B, p38, and stress-activated protein kinase mitogen-activated protein kinase signaling pathways regulate proinflammatory cytokines and apoptosis in human placental explants in response to oxidative stress: Effects of antioxidant vitamins. Am. J. Pathol. 2007, 170, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.T.; Wei, W.; Ma, B.; Xu, Y.; Liu, W.J.; Wang, Y.; Lv, K.Y.; Tang, H.T.; Wei, D.; Xia, Z.F. Activation of p38 MAPK by reactive oxygen species is essential in a rat model of stress-induced gastric mucosal injury. J. Immunol. 2007, 179, 7808–7819. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.T.; Shih, R.H.; Lin, C.C.; Chen, J.T.; Yang, C.M. Role of TLR4/NADPH oxidase/ROS-activated p38 MAPK in VCAM-1 expression induced by lipopolysaccharide in human renal mesangial cells. Cell Commun. Signal. 2012, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Papaconstantinou, J. p38 Mitogen activated protein kinase (MAPK): A new therapeutic target for reducing the risk of adverse pregnancy outcomes. Expert. Opin. Ther. Targets 2016, 20, 1397–1412. [Google Scholar] [CrossRef] [PubMed]
- Goldman, B.; Radnaa, E.; Kechichian, T.; Menon, R. Silencing P38 MAPK reduces cellular senescence in human fetal chorion trophoblast cells. Am. J. Reprod. Immunol. 2023, 89, e13648. [Google Scholar] [CrossRef] [PubMed]
- Sheller-Miller, S.; Richardson, L.; Martin, L.; Jin, J.; Menon, R. Systematic review of p38 mitogen-activated kinase and its functional role in reproductive tissues. Am. J. Reprod. Immunol. 2018, 80, e13047. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.; Choe, J. Prostaglandin E2 stimulates COX-2 expression via mitogen-activated protein kinase p38 but not ERK in human follicular dendritic cell-like cells. BMC Immunol. 2020, 21, 20. [Google Scholar] [CrossRef] [PubMed]
- Parente, R.; Trifirò, E.; Cuozzo, F.; Valia, S.; Cirone, M.; Di Renzo, L. Cyclooxygenase-2 is induced by p38 MAPK and promotes cell survival. Oncol. Rep. 2013, 29, 1999–2004. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.M.; Xu, W.M.; Lin, J.C.; Mo, L.Q.; Hua, X.X.; Chen, P.X.; Wu, K.; Zheng, D.D.; Feng, J.Q. Activation of the p38 MAPK/NF-κB pathway contributes to doxorubicin-induced inflammation and cytotoxicity in H9c2 cardiac cells. Mol. Med. Rep. 2013, 8, 603–608. [Google Scholar] [CrossRef]
- Richardson, L.; Dixon, C.L.; Aguilera-Aguirre, L.; Menon, R. Oxidative stress-induced TGF-beta/TAB1-mediated p38MAPK activation in human amnion epithelial cells. Biol. Reprod. 2018, 99, 1100–1112. [Google Scholar] [CrossRef] [PubMed]
- Gotsch, F.; Romero, R.; Chaiworapongsa, T.; Erez, O.; Vaisbuch, E.; Espinoza, J.; Kusanovic, J.P.; Mittal, P.; Mazaki-Tovi, S.; Kim, C.J.; et al. Evidence of the involvement of caspase-1 under physiologic and pathologic cellular stress during human pregnancy: A link between the inflammasome and parturition. J. Matern. Fetal Neonatal Med. 2008, 21, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Miller, D.; Leng, Y.; Panaitescu, B.; Silva, P.; Faro, J.; Alhousseini, A.; Gill, N.; et al. The immunophenotype of amniotic fluid leukocytes in normal and complicated pregnancies. Am. J. Reprod. Immunol. 2018, 79, e12827. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Parvizi, S.T.; Oyarzun, E.; Mazor, M.; Wu, Y.K.; Avila, C.; Athanassiadis, A.P.; Mitchell, M.D. Amniotic fluid interleukin-1 in spontaneous labor at term. J. Reprod. Med. 1990, 35, 235–238. [Google Scholar] [PubMed]
- Gomez-Lopez, N.; Romero, R.; Panaitescu, B.; Miller, D.; Zou, C.; Gudicha, D.W.; Tarca, A.L.; Para, R.; Pacora, P.; Hassan, S.S.; et al. Gasdermin D: In vivo evidence of pyroptosis in spontaneous labor at term. J. Matern. Fetal Neonatal Med. 2021, 34, 569–579. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Motomura, K.; Miller, D.; Garcia-Flores, V.; Galaz, J.; Romero, R. Inflammasomes: Their Role in Normal and Complicated Pregnancies. J. Immunol. 2019, 203, 2757–2769. [Google Scholar] [CrossRef]
- Panaitescu, B.; Romero, R.; Gomez-Lopez, N.; Xu, Y.; Leng, Y.; Maymon, E.; Pacora, P.; Erez, O.; Yeo, L.; Hassan, S.S.; et al. In vivo evidence of inflammasome activation during spontaneous labor at term. J. Matern. Fetal Neonatal Med. 2019, 32, 1978–1991. [Google Scholar] [CrossRef] [PubMed]
- Stutz, A.; Horvath, G.L.; Monks, B.G.; Latz, E. ASC speck formation as a readout for inflammasome activation. Methods Mol. Biol. 2013, 1040, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Garcia-Flores, V.; Leng, Y.; Panaitescu, B.; Miller, D.; Abrahams, V.M.; Hassan, S.S. Inflammasome assembly in the chorioamniotic membranes during spontaneous labor at term. Am. J. Reprod. Immunol. 2017, 77, e12648. [Google Scholar] [CrossRef] [PubMed]
- Motomura, K.; Romero, R.; Galaz, J.; Tao, L.; Garcia-Flores, V.; Xu, Y.; Done, B.; Arenas-Hernandez, M.; Miller, D.; Gutierrez-Contreras, P.; et al. Fetal and maternal NLRP3 signaling is required for preterm labor and birth. JCI Insight 2022, 7, e158238. [Google Scholar] [CrossRef] [PubMed]
- Colon-Caraballo, M.; Lee, N.; Nallasamy, S.; Myers, K.; Hudson, D.; Iozzo, R.V.; Mahendroo, M. Novel regulatory roles of small leucine-rich proteoglycans in remodeling of the uterine cervix in pregnancy. Matrix Biol. 2022, 105, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Akgul, Y.; Word, R.A.; Ensign, L.M.; Yamaguchi, Y.; Lydon, J.; Hanes, J.; Mahendroo, M. Hyaluronan in cervical epithelia protects against infection-mediated preterm birth. J. Clin. Investig. 2014, 124, 5481–5489. [Google Scholar] [CrossRef] [PubMed]
- Pacora, P.; Romero, R.; Maymon, E.; Gervasi, M.T.; Gomez, R.; Edwin, S.S.; Yoon, B.H. Participation of the novel cytokine interleukin 18 in the host response to intra-amniotic infection. Am. J. Obstet. Gynecol. 2000, 183, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.N.; Rao, L.; Sha, Y.; Abdel Fattah, E.; Hyser, J.; Eissa, N.T. p38 MAPK Activity Is Required to Prevent Hyperactivation of NLRP3 Inflammasome. J. Immunol. 2021, 207, 661–670. [Google Scholar] [CrossRef]
- Li, D.; Ren, W.; Jiang, Z.; Zhu, L. Regulation of the NLRP3 inflammasome and macrophage pyroptosis by the p38 MAPK signaling pathway in a mouse model of acute lung injury. Mol. Med. Rep. 2018, 18, 4399–4409. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best. Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Hayyan, M.; Hashim, M.A.; AlNashef, I.M. Superoxide Ion: Generation and Chemical Implications. Chem. Rev. 2016, 116, 3029–3085. [Google Scholar] [CrossRef]
- Muller, F. The nature and mechanism of superoxide production by the electron transport chain: Its relevance to aging. J. Am. Aging Assoc. 2000, 23, 227–253. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.C. Antimicrobial actions of reactive oxygen species. mBio 2011, 2, e00141-11. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L.; Yeo, D. Oxidative stress: An evolving definition. Fac. Rev. 2021, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Ruder, E.H.; Hartman, T.J.; Goldman, M.B. Impact of oxidative stress on female fertility. Curr. Opin. Obstet. Gynecol. 2009, 21, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Al-Gubory, K.H.; Fowler, P.A.; Garrel, C. The roles of cellular reactive oxygen species, oxidative stress and antioxidants in pregnancy outcomes. Int. J. Biochem. Cell Biol. 2010, 42, 1634–1650. [Google Scholar] [CrossRef] [PubMed]
- Raijmakers, M.T.; Burton, G.J.; Jauniaux, E.; Seed, P.T.; Peters, W.H.; Steegers, E.A.; Poston, L. Placental NAD(P)H oxidase mediated superoxide generation in early pregnancy. Placenta 2006, 27, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Schofield, J.H.; Schafer, Z.T. Mitochondrial Reactive Oxygen Species and Mitophagy: A Complex and Nuanced Relationship. Antioxid. Redox Signal. 2021, 34, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Vermot, A.; Petit-Härtlein, I.; Smith, S.M.E.; Fieschi, F. NADPH Oxidases (NOX): An Overview from Discovery, Molecular Mechanisms to Physiology and Pathology. Antioxidants 2021, 10, 890. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.C.; Andriantsitohaina, R. Reactive nitrogen species: Molecular mechanisms and potential significance in health and disease. Antioxid. Redox Signal. 2009, 11, 669–702. [Google Scholar] [CrossRef] [PubMed]
- Jha, N.; Ryu, J.J.; Choi, E.H.; Kaushik, N.K. Generation and Role of Reactive Oxygen and Nitrogen Species Induced by Plasma, Lasers, Chemical Agents, and Other Systems in Dentistry. Oxid. Med. Cell. Longev. 2017, 2017, 7542540. [Google Scholar] [CrossRef] [PubMed]
- Cindrova-Davies, T.; Yung, H.W.; Johns, J.; Spasic-Boskovic, O.; Korolchuk, S.; Jauniaux, E.; Burton, G.J.; Charnock-Jones, D.S. Oxidative stress, gene expression, and protein changes induced in the human placenta during labor. Am. J. Pathol. 2007, 171, 1168–1179. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Alfadda, A.A.; Sallam, R.M. Reactive oxygen species in health and disease. J. Biomed. Biotechnol. 2012, 2012, 936486. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Roychoudhury, S. Reactive Oxygen Species in the Reproductive System: Sources and Physiological Roles. Adv. Exp. Med. Biol. 2022, 1358, 9–40. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Richardson, L.; Sheller-Miller, S.; Zhong, N.; Menon, R. Oxidative stress induces p38MAPK-dependent senescence in the feto-maternal interface cells. Placenta 2018, 67, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Keshari, R.S.; Verma, A.; Barthwal, M.K.; Dikshit, M. Reactive oxygen species-induced activation of ERK and p38 MAPK mediates PMA-induced NETs release from human neutrophils. J. Cell. Biochem. 2013, 114, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Mulero, M.C.; Huxford, T.; Ghosh, G. NF-κB, IκB, and IKK: Integral Components of Immune System Signaling. Adv. Exp. Med. Biol. 2019, 1172, 207–226. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Nakajima, S.; Kitamura, M. Bidirectional regulation of NF-κB by reactive oxygen species: A role of unfolded protein response. Free Radic. Biol. Med. 2013, 65, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS Crosstalk in Inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Castro, J.; Florido, J.; Kajarabille, N.; Prados, S.; de Paco, C.; Ocon, O.; Pulido-Moran, M.; Ochoa, J.J. A new approach to oxidative stress and inflammatory signaling during labour in healthy mothers and neonates. Oxid. Med. Cell. Longev. 2015, 2015, 178536. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Elner, S.G.; Bian, Z.M.; Till, G.O.; Petty, H.R.; Elner, V.M. Pro-inflammatory cytokines increase reactive oxygen species through mitochondria and NADPH oxidase in cultured RPE cells. Exp. Eye Res. 2007, 85, 462–472. [Google Scholar] [CrossRef]
- Qayyum, N.; Haseeb, M.; Kim, M.S.; Choi, S. Role of Thioredoxin-Interacting Protein in Diseases and Its Therapeutic Outlook. Int. J. Mol. Sci. 2021, 22, 2754. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Chen, F.; Yin, Q.; Wang, D.; Han, W.; Zhang, Y. Reactive Oxygen Species Interact with NLRP3 Inflammasomes and Are Involved in the Inflammation of Sepsis: From Mechanism to Treatment of Progression. Front. Physiol. 2020, 11, 571810. [Google Scholar] [CrossRef] [PubMed]
- Dominic, A.; Le, N.T.; Takahashi, M. Loop between NLRP3 Inflammasome and Reactive Oxygen Species. Antioxid. Redox Signal. 2022, 36, 784–796. [Google Scholar] [CrossRef]
- Maynard, S.; Schurman, S.H.; Harboe, C.; de Souza-Pinto, N.C.; Bohr, V.A. Base excision repair of oxidative DNA damage and association with cancer and aging. Carcinogenesis 2009, 30, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Rojo, C.; Rodríguez-Orozco, A.R. Importance of oxidative damage on the electron transport chain for the rational use of mitochondria-targeted antioxidants. Mini Rev. Med. Chem. 2011, 11, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Nakahira, K.; Haspel, J.A.; Rathinam, V.A.; Lee, S.J.; Dolinay, T.; Lam, H.C.; Englert, J.A.; Rabinovitch, M.; Cernadas, M.; Kim, H.P.; et al. Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome. Nat. Immunol. 2011, 12, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, Y.; Inazumi, T.; Tsuchiya, S. Roles of prostaglandin receptors in female reproduction. J. Biochem. 2015, 157, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Arrowsmith, S.; Wray, S. Oxytocin: Its mechanism of action and receptor signalling in the myometrium. J. Neuroendocrinol. 2014, 26, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lopez, N.; Romero, R.; Garcia-Flores, V.; Leng, Y.; Miller, D.; Hassan, S.S.; Hsu, C.D.; Panaitescu, B. Inhibition of the NLRP3 inflammasome can prevent sterile intra-amniotic inflammation, preterm labor/birth, and adverse neonatal outcomes†. Biol. Reprod. 2019, 100, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Faro, J.; Romero, R.; Schwenkel, G.; Garcia-Flores, V.; Arenas-Hernandez, M.; Leng, Y.; Xu, Y.; Miller, D.; Hassan, S.S.; Gomez-Lopez, N. Intra-amniotic inflammation induces preterm birth by activating the NLRP3 inflammasome†. Biol. Reprod. 2019, 100, 1290–1305. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, C.; Swartzwelter, B.; Gamboni, F.; Neff, C.P.; Richter, K.; Azam, T.; Carta, S.; Tengesdal, I.; Nemkov, T.; D’Alessandro, A.; et al. OLT1177, a β-sulfonyl nitrile compound, safe in humans, inhibits the NLRP3 inflammasome and reverses the metabolic cost of inflammation. Proc. Natl. Acad. Sci. USA 2018, 115, E1530–E1539. [Google Scholar] [CrossRef]
Figure 1.
Scheme showing the overall course of biochemical and molecular pathways occurring during cervical ripening; ROS—reactive oxygen species; IL-1—interleukin-1; NF-kB—nuclear factor kappa-B; IL-18—interleukin-18; p38MAPK—p38 mitogen-activated protein kinase; PGs—prostaglandins; NO—nitric oxide; MMPs—metalloproteinases.
Figure 1.
Scheme showing the overall course of biochemical and molecular pathways occurring during cervical ripening; ROS—reactive oxygen species; IL-1—interleukin-1; NF-kB—nuclear factor kappa-B; IL-18—interleukin-18; p38MAPK—p38 mitogen-activated protein kinase; PGs—prostaglandins; NO—nitric oxide; MMPs—metalloproteinases.

Figure 2.
Diagram showing possible role of NLPR3 inflammasome in cervical ripening; NF-kB—nuclear factor kappa-B; ROS—reactive oxygen species; IL-1—interleukin-1; IL-18—interleukin-18; p38MAPK—p38 mitogen-activated protein kinase; PGs—prostaglandins; NO—nitric oxide; MMPs—metalloproteinases; COX-2—cyclooxygenase-2; TNF-α—tumor necrosis factor-α; HAS—hyaluronan synthase; HA—hyaluronan.
Figure 2.
Diagram showing possible role of NLPR3 inflammasome in cervical ripening; NF-kB—nuclear factor kappa-B; ROS—reactive oxygen species; IL-1—interleukin-1; IL-18—interleukin-18; p38MAPK—p38 mitogen-activated protein kinase; PGs—prostaglandins; NO—nitric oxide; MMPs—metalloproteinases; COX-2—cyclooxygenase-2; TNF-α—tumor necrosis factor-α; HAS—hyaluronan synthase; HA—hyaluronan.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Flis, W.; Socha, M.W. The Role of the NLRP3 Inflammasome in the Molecular and Biochemical Mechanisms of Cervical Ripening: A Comprehensive Review. Cells 2024, 13, 600. https://doi.org/10.3390/cells13070600
AMA Style
Flis W, Socha MW. The Role of the NLRP3 Inflammasome in the Molecular and Biochemical Mechanisms of Cervical Ripening: A Comprehensive Review. Cells. 2024; 13(7):600. https://doi.org/10.3390/cells13070600
Chicago/Turabian StyleFlis, Wojciech, and Maciej W. Socha. 2024. "The Role of the NLRP3 Inflammasome in the Molecular and Biochemical Mechanisms of Cervical Ripening: A Comprehensive Review" Cells 13, no. 7: 600. https://doi.org/10.3390/cells13070600
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.