Functional Traits of Plant Species Suitable for Revegetation of Landfill Waste from Nickel Smelter


1
Department of Soil Science, Faculty of Natural Sciences, Comenius University, Mlynská dolina, Ilkovičova 6, 842 15 Bratislava, Slovakia
2
Institute of Hydrology SAS, Dúbravská cesta 9, 841 04 Bratislava, Slovakia
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(2), 658; https://doi.org/10.3390/app11020658
Submission received: 7 December 2020
/
Revised: 27 December 2020
/
Accepted: 8 January 2021
/
Published: 12 January 2021
(This article belongs to the Section Environmental Sciences)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
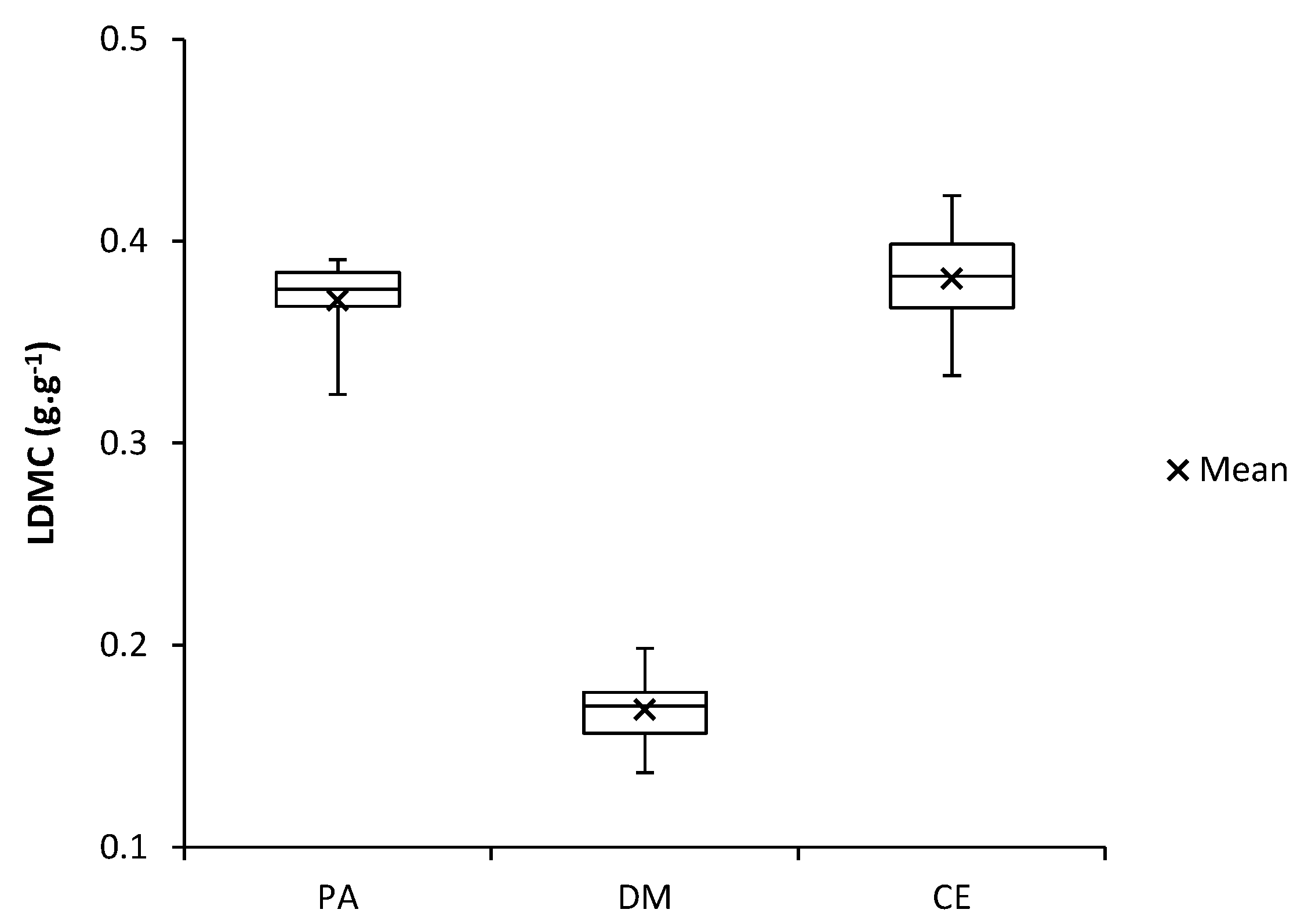{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The landfill waste of leached ore residue represents a serious environmental risk and may also negatively affect the appearance, growth and development of vegetation. Here we focused on the evaluation of functional traits of selected plant species Populus alba, Calamagrostis epigejos, and Diplotaxis muralis growing in an unfavourable environment. We determined different adaptive strategies of selected species to extreme conditions. For Diplotaxis muralis the highest values of the leaf dry matter content (LDMC) and the lowest values of the specific leaf area (SLA) were determined, while for Calamagrostis epigejos these two traits correlated in opposite directions. Populus alba reached the lowest value of the water saturation deficit (WSD), suggesting that this species was most affected by soil water deficiency. The leaf water content (LWC) correlated negatively with the LDMC and positively with the SLA (narrow leaf blade). Although each plant species belongs to a different strategic group (therophyte, hemicryptophyte and phanerophyte in the juvenile stage), they are all very plastic and therefore suitable for remediation. Despite the unfavourable conditions, selected plant species were able to adapt to poor conditions and form more or less vital populations, which indicate the revegetation as a key measure for remediation of landfill waste from nickel smelter.
1. Introduction
Industrial activities leave unwanted traces in the surrounding territory and have a severe impact on the environment. As is well known, industrial and mining wastes with a high content of heavy metals pose a risk to the environment and have a demonstrable toxic effect on vegetation and soil microorganisms and their life processes [1,2,3]. These environmental burdens represent a very negative factor influencing the functional spatial structure of the landscape. There is not only contaminated groundwater around the landfill but also a high level of air pollution with toxic dust. [4]. Recently, various methods have been used worldwide for the remediation of similarly contaminated sites: isolation and stabilization of geochemically transformed elements [5,6,7], using natural alkaline material [8], covering of mine tailings by using inert materials [9], soil washing [10], electrokinetic remediation [11], microbiological methods [12] and many others. These technologies can be successful but generally require several specific processes, which take a long time. They are often expensive and can be practically used only in small areas [13]. Therefore, there is currently an increasing emphasis on the use of various phytotechnologies, including phytoextraction, phytodegradation, rhizofiltration, phytostabilization, phytovolatilization or phytocapping [1,13,14,15]. Revegetation is a low-cost measure based on the use of naturally occurring plants to cover mining sites to not allow a release of heavy metals from mining waste into surrounding soil, groundwater and air by creating natural surface biomass layer as well as a branched root system. Thanks to this plant cover, water and wind erosion are also prevented, rhizosphere properties can be improved and thus the landfill can be gradually revitalized [13].
The investigated site was the landfill waste of black mud near the town of Sereď (SW Slovakia), which is a source of heavy metal contamination. The landfill environment is unfavourable for plant growth. Landfill material is not soil in the real sense; it is industrial waste that has poor physical properties without developed soil structure. Due to its black colour, the landfill surface heats up and dries out very quickly. The material is strongly alkaline (pH 8.5) and the organic content is very low 0.05%. The C:N ratio is 0.74:0.05. Total content of Fe2O3 is 78% and Al2O3 3.27%. Content of chromium is the highest 24,300 mg/kg, nickel 2920 mg/kg, zinc 300 mg/kg and copper 79 mg/kg. [16]. It does not contain enough biogenic elements for the proper plant growth and nutrition. Greening of the landfill waste can be considered as one of the most natural, and in this case, the most appropriate remediation methods.
This work was aimed to characterise native plant species that have been able to adapt in an extreme environment unfavourable for plant growth. The functional traits of three plant species Populus alba, Calamagrostis epigejos and Diplotaxis muralis were evaluated, which reflect the plant’s response to the habitat and a way of adapting to environmental conditions.
1.1. Site Characterisation
In the second half of the last century, nickel ore was processed near the town of Sereď (SW Slovakia, Figure 1), where the landfill waste of black mud (waste from the colour metallurgy of nickel and cobalt) is a direct evidence of this activity. The tailing is 45 m high, 800 m long and 550 m wide (Figure 2). In the south, west and east, it is in contact with agricultural land and represents a significant anthropogenic barrier [16]. It is a distinct anthropogenic form of relief with a typical convex unit (Figure 2). The site is noncombustible and has a distinctive shape of a tabular terrace, which was formed by the gradual deposition of waste layers—black nickel mud. It has steep slopes of up to 45 degrees (anthropogenic table mountain). According to a field survey by Michaeli et al. [4], the landfill waste has high permeability; therefore, rainwater quickly penetrates the lower layers, and the remaining water evaporates quickly since the black nickel mud strongly absorbs sunlight and thus rapidly overheats.
After the closure of the smelting factory in Sereď, the landfill waste was formed by reclamation processes, geomorphological processes and processes from anthropogenic activities (mining of black mud). The landfill soil can be classified as technosol developed form a substrate of technogenic origin [16].
1.2. Characteristics of Investigated Plant Species
To determine the functional traits, three plant species were selected, which represent different life forms—therophyte, hemicryptophyte and phanerophyte (in the juvenile stage).
Populus alba (Figure 3, white poplar—phanerophyte [17] in the juvenile stage) is a fast-growing tree with deciduous leaves, which can grow to a height of 20 to 35 m. Dark green, lobed leaves, the underside is slightly felted, coloured white. It is widespread in Central, Southern and Eastern Europe, across Siberia to the Yenisei River, in Asia to the western Himalayas. It was introduced to Micronesia, Australia and New Zealand and is also spreading invasively in the Canary Islands. It is a tree of floodplain forests; it occurs in coastal shrubs and on river alluvium. In the soft meadow, it forms communities with Salix alba (white willow) and Populus nigra (black poplar). White poplar is a light-loving tree, only younger individuals tolerate weaker shading. It tolerates changes in water regime very well and is also resistant to long-term floods. It is secondary to quarries in sandpits, brickyards, embankments, landfills, mining heaps and similar places with exposed surfaces [18]. In Slovakia, it is widespread in the lowlands [19].
Calamagrostis epigejos (Figure 4, bush grass—hemicryptophyte [17] is a perennial grey-green bushy grass that has strong creeping subsoils with long shoots. It inhabits mainly sparse forests, log cabins, quarries, neglected pastures and ruderal areas. In pastures and meadows, this species is invasive and suppresses various meadow communities. Bush grass is an ecologically plastic and can colonize multiple substrates. It is widespread in Europe, North Asia, and secondarily in North America and South Africa. It inhabits the whole territory of Slovakia [20].
Diplotaxis muralis (Figure 5, annual wall-rocket—therophyte [17]) is an annual, biennial, rarely perennial herb, 15–50 cm tall. It inhabits mainly rubbles, fields, fallows and vineyards; it also occurs along roads, on landfills, ruderalized lawns, composts and various anthropogenic habitats. It occurs mostly on clayey or gravelly, nutrient-rich soils [21]. It is originally widespread throughout Central and Southern Europe (up to Northwest Africa), secondarily it occurs in Northern Europe, in parts of Russia and northern China, in South Africa, as well as in Australia and Tasmania, New Zealand, New Mexico, in almost the entire United States and southern Canada, and Central and South America [22]. In Slovakia, it is a common species, especially in warmer areas and in habitats with mild to medium soil moisture.
1.3. Functional Traits
Functional traits are defined as morpho-physio-phenological traits, which impact the fitness of plant indirectly via their effects on growth, reproduction and survival [23]. Some plants have the ability to adapt to changes in the environment; this feature is called phenotypic plasticity. Plasticity may vary from species to species, some species are unable to adapt to changes in the environment, but some have evolved, and they are able to respond positively to changes, i.e., changes in soil properties due to contamination, changes in water management, climatic changes, etc. [24].
Functional traits can be divided into simple traits—soft traits, which can be quantified relatively quickly and easily, and hard traits, which are more difficult to quantify. Soft traits include the water and dry matter content of leaves, the specific leaf area, the number of seeds, the relative length of the roots, the bark thickness or the height of the plant.
1.3.1. Specific Leaf Area (SLA)
It represents the area of one side of a fresh leaf divided by its dry matter. It is expressed in cm2⋅g−1. For example, species with higher SLAs (less carbon invested per leaf area) achieve higher specific growth rates (RGRs), which is an essential criterion for assessing biomass production, to compare plant species performance as well or to observe different impacts on plants. Plants with higher SLA values have thinner leaves and higher chlorophyll concentration per unit weight [25,26].
Further studies also found a positive correlation between SLA and the rate of net photosynthesis. SLA has also proven to be a key parameter that allows a plant to expose a large area of leaves to CO2 and light, and thus use them efficiently. Species found in environments with enough nutrients, water, light, etc., generally have higher SLA values than species living in a resource-poor environment. Low values tend to correspond to long leaf life and relatively high contributions to the defence mechanisms of plant species [27].
1.3.2. Leaf Dry Matter Content (LDMC)
It is expressed in units of g⋅g−1 and represents the dry weight of the leaves divided by its fresh weight. In this case, the fresh weight corresponds to 100%. The remainder is water in the leaves, and therefore the water content of the leaves (LWC) can be derived as 100%—LDMC or 1—LDMC [25,28].
LDMC is closely related to leaf tissue density. There is a negative correlation with the relative potential growth rate but a positive correlation with leaf life. However, the resulting benefits are likely to be less than those of the SLA. Leaves with a high LDMC value are relatively stiff so that they may be more resistant to physical risks (hail, wind, herbivores). Leaves with low LDMC are associated with a considerably disturbed environment. The dry matter content of the leaves is more useful and can provide more information than the SLA in cases where it is difficult to measure the leaf area [27].
1.3.3. Water Saturation Deficit (WSD)
Water saturation deficit means the amount of water that is missing (or part of it) in the plant until it is fully saturated; in other words, the difference between the water content at maximum saturation and its content at a given moment. It is expressed as a percentage of the maximum water content. A water deficit occurs when the water balance is disrupted, and the plant releases more water than it receives [25].
Water balance describes the correlation between water loss and water uptake. When the plant is optimally saturated with water, an optimal water balance occurs. Temporary or permanent decrease of water saturation of the plant with negative influence and effects occurs after depletion of physiologically available water. Water saturation of the plant occurs at an active water balance, and thus a water deficit occurs at a negative water balance. A sublethal water deficit is typical water stress and thus a borderline case of the water deficit. It expresses the value of WSD when 5–10% of leaf area is already irreversibly damaged [29].
1.3.4. Leaf Water Content (LWC)
In the physiological processes of plants, the water content of the leaves is an essential indicator of photosynthesis, transpiration or respiration [30].
The water regime of the plant is crucial for the proper functioning of the cycle of substances in plant tissues. The water content in the leaves is thus an essential prerequisite for plant growth, especially to prevent drought stress. The water content at leaf level is derived from the difference between the weight of freshly harvested leaves and their dry weight or the 1-LDMC relationship [31].
2. Materials and Methods
2.1. Field Work and Collection of Leaves
Regarding the aims of the work, samples of plant material were randomly taken from an area of approximately 20 × 20 m2 (Figure 2). Three dominant species were sampled: Populus alba, Calamagrostis epigejos and Diplotaxis muralis. Ten leaf samples, for Populus alba in the juvenile stage, were taken in July and September from ten individuals per genus that were not damaged by external negative influences (herbivores, pathogens). We removed the leaves, which were vital, healthy, developed, together with the stem, in the place of natural separation at the leaf fall.
2.2. Functional Traits
The measurement of the functional traits of the plants was in accordance with the procedure described in Cornelissen et al. [27].
The leaf characteristics of the plants that have been determined include:
- FM—fresh leaf weight [g]
- SM—saturated leaf weight [g]
- DM—dry weight of the leaf [g]
- LA—leaf area [cm2]
Using these values, the functional traits were calculated as
- Specific leaf area: SLA = LA/DM [cm2·g−1]
- Leaf dry matter content: LDMC = DM/FM [g·g−1]
- Water saturation deficit: WSD = (SM-FM)/SM [%]
- Leaf water content: LWC = 1-LDMC [g·g−1]
2.3. Statistical Analysis
Box plots in the graphs show the variability of individual functional leaf traits (SLA, LDMC, WSD, LWC) between 3 selected species. Boxplots are a standardized way of displaying the distribution of data based on a five-number summary: minimum, first quartile, median, third quartile and maximum. The edges of the box represent the upper and lower quartiles (values in the range of 25–75% of the total range of values).
Pearson’s correlation coefficients (r) quantified the linear relationships between functional traits. Correlation relationships were classified as: small dependence for r ≤ 0.4, slight dependence for 0.4 < r ≤ 0.8 and strong dependence for r > 0.8.
3. Results
3.1. Specific Leaf Area
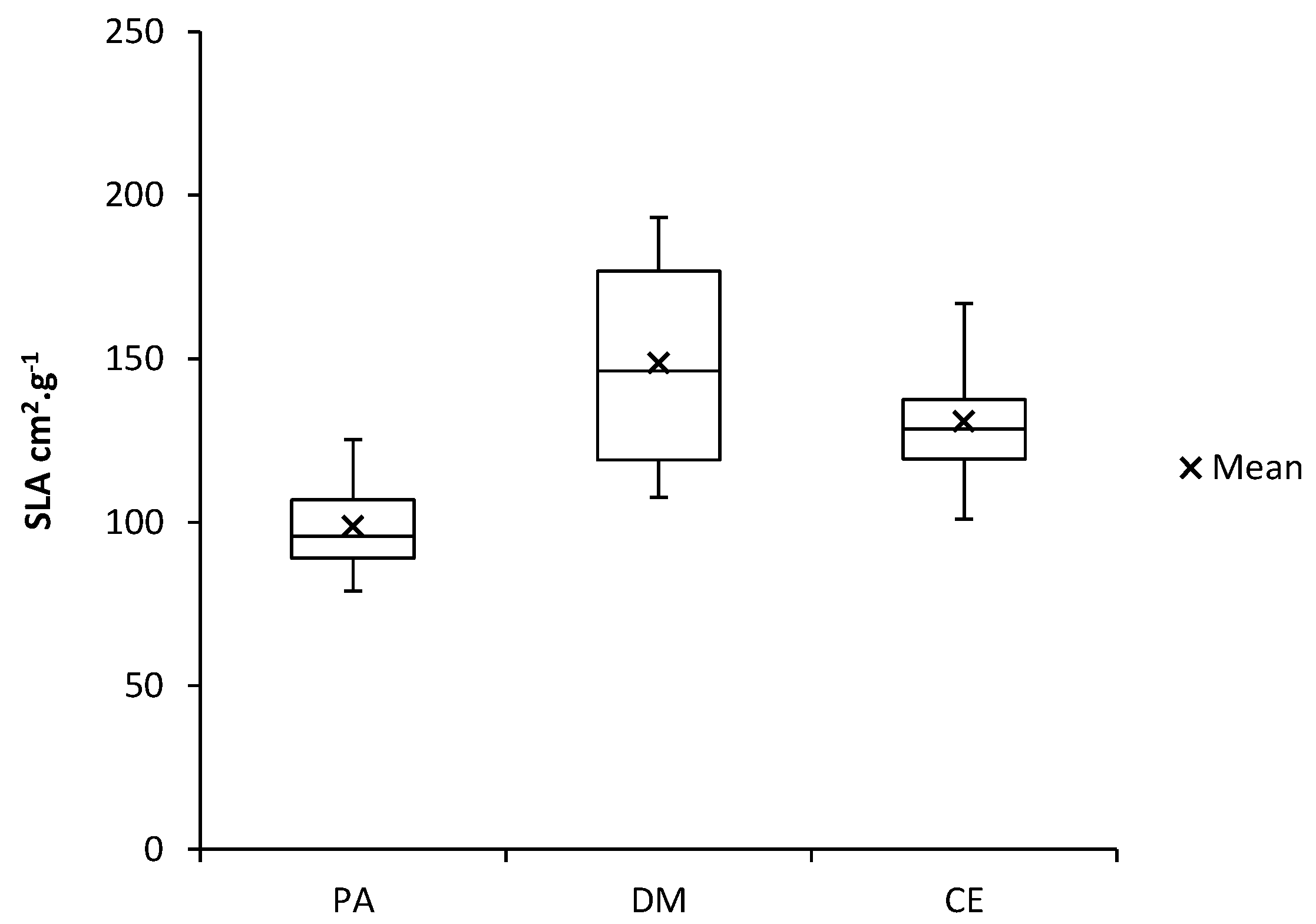
The Diplotaxis muralis had the highest average values of specific leaf area (SLA) of 148.7 cm2·g−1, the Calamagrostis epigejos followed with an average value of 130.77 cm2·g−1 and the lowest values were found for the Populus alba 98.7 cm2·g−1 (Figure 6).
In terms of functional groups, according to the values shown in Figure 6, we could include the species Calamagrostis epigejos and Diplotaxis muralis in one group and the Populus alba in another group. This probably reflects different life strategies, various adaptations of species and acclimatization of their organs to given environmental conditions.
3.2. Leaf Dry Matter Content
The differences in values of the dry matter content in the leaves (LDMC) between selected species are shown in Figure 7. The LDMC values are the lowest for the Diplotaxis muralis and, conversely, the LDMC values are much higher for the other two species Populus alba and Calamagrostis epigejos.
The LDMC values are the lowest for the Diplotaxis muralis and, conversely, the LDMC values are much higher for the other two species Populus alba and Calamagrostis epigejos. Therefore, we could include Populus alba and Calamagrostis epigejos in one functional group, which indicate their similar mechanism of adaptation to extreme properties of soil on the landfill.
We found the highest average values of the LDMC in the Populus alba (0.37 g·g−1) and Calamagrostis epigejos (0.38 g·g−1), and the lowest values for the Diplotaxis muralis (0.17 g·g−1).
3.3. Water Saturation Deficit
Figure 8 shows the differences between water saturation deficits in the selected species. The highest average value was determined in the Populus alba with the WSD of 33.2%; for the Diplotaxis muralis it was 20.8% and for the Calamagrostis epigejos the lowest average value was 8.1%. The Calamagrostis epigejos probably has the best strategy for water management under stress.
3.4. Leaf Water Content
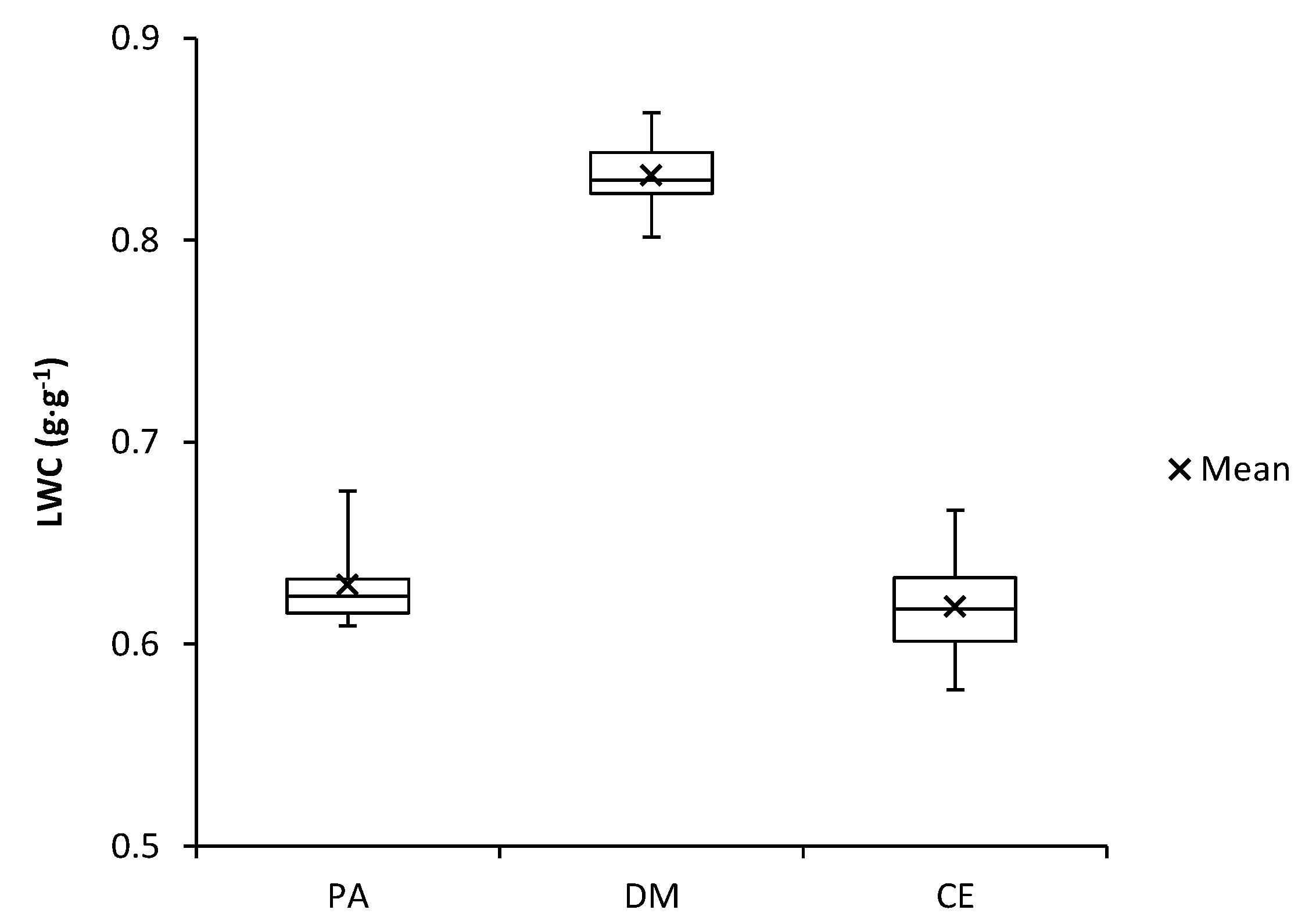
The measured values of the LWC (Figure 9) showed that the Diplotaxis muralis is, despite the unfavourable conditions in the landfill, able to work with water most effectively and keep it in tissues. The highest measured value for the Diplotaxis muralis was 0.86 g·g−1. The differences between the studied species were not too high, and the measured values indicate that the Populus alba and Calamagrostis epigejos are also able to make full use of their water potential. The highest measured value for the Populus alba was 0.68 g·g−1 and for the Calamagrostis epigejos 0.67 g·g−1.
3.5. Correlation between SLA and LWC
Based on the values of individual functional traits, we decided to determine whether the water content in the leaves (LWC) affects the specific leaf area (SLA). In the correlation diagram (Figure 10), the relationship between the SLA and LWC is plotted, which is significant for the Populus alba with r = 0.716.
For the other two species, the dependence was not statistically significant. Low correlation between SLA and LWC for the Diplotaxis muralis (r = 0.386) is most likely caused by its adaptation to the lack of water in the habitat. The plant has scleromorphic leaves and can create an extensive root system [32].
The correlation coefficients in Figure 10 may indicate either different plant forms: the Populus alba is juvenile, the Calamagrostis epigejos is grass and Diplotaxis muralis is a herb; or it can mean a favourable leaf morphology as in the case of Populus alba, which is characterised with a larger leaf area and with the presence of hairs on the leaf surface, and thus it can better manage water.
4. Discussion
4.1. Functional Traits and Ecological Plasticity
Low SLA values are typical for species inhabiting environments with low nutrient content, lack of water and light [27]. Our results confirmed low SLA values, mainly for the type of Populus alba. However, it should be noted that all selected species occurred in the area outside the shade with enough light, which is an essential factor for SLA, because its values are mainly related to the photosynthetic activity of the leaves. Despite the lowest achieved SLA values, we can agree with the findings of Rafati et al. [33] that the Populus alba species is able to colonize contaminated substrates and grow even in extreme conditions. In terms of its ecological plasticity, Populus alba showed rapid growth, high biomass production, easy seed dispersal, fast attachment and root system development.
We can classify Calamagrostis epigejos among C-strategists at our site. In addition, according to Mitrovica et al. [32], Calamagrostis epigejos has a high growth potential, which could be used especially in disturbed habitats for shortening succession series and faster reclamation.
The resulting SLA values for Diplotaxis muralis indicate that this species is the best of the selected species adapted to the conditions of the habitat. This is probably due to the fact that this species can be included among the R-strategists which enter the habitats in the early stages of succession, have the high reproductive capacity and rapid germination as well as rapid population growth with short life cycle and short vegetation phase.
Higher water content in the leaves and a narrower leaf blade contribute to a higher SLA [34,35]. Plants with higher SLA flexibly respond to changes in resource availability in the soil [36], are more productive [37] but more prone to damage herbivores [38]. Cingolani et al. [39] showed that productivity-related traits (SLA, LDMC) strongly reflect the soil moisture gradient.
The LDMC reflects the availability of nutrients and water in the soil. Based on the obtained results, we can state that the Diplotaxis muralis species is well adapted to adverse conditions, as selected species were investigated in habitats with low soil moisture and nutrient content but with a high content of heavy metals. Its leaves are stiff, which is also a manifestation of its resistance to adverse external influences and its ability to use nutrients efficiently. Thus, it can survive long periods without water supply. For Populus alba and Calamagrostis epigejos species, regardless of their higher LDMC values, we assume that they are also resistant to adverse habitat conditions. In the case of the Populus alba, even though it requires higher humidity, it can also occur in water-deficient habitats. Calamagrostis epigejos is an ecologically plastic species which can occur in adverse habitats. We believe that this species was able to adapt well to extreme conditions in the habitat.
The LDMC and SLA are highly correlated characteristics that reflect different plant adaptations. Each of these properties may indicate different functional strategies, where the SLA is mainly related to the light availability and photosynthetic activity, and the LDMC indicates the water and nutrient availability in soil.
In terms of different adaptations and life strategies, we can divide selected species into two functional groups. Diplotaxis muralis belongs to the first functional group with lower LDMC values. We assume that it is well adapted to the given habitat and can effectively use available nutrients and water. Based on the higher values (dry matter content in the leaves) of the Populus alba and Calamagrostis epigejos species, we believe that they are more stressed by the adverse effects of the habitat and thus handle the obtained nutrients and water more gently. Therefore, in terms of the life strategy and the use of resources, we classify Populus alba and Calamagrostis epigejos into the second functional group of the examined habitat.
The highest values of WSD were found in Populus alba. Its highest value reached up to 83.7%, which is the limit value of the water deficit [40]. The water deficit threshold, or sublethal water deficit, is a condition in which irreversible leaf damage occurs. Sublethal water deficit is achieved at values in the range of 70–80% WSD. In our case, one sample of Populus alba reached the value of the sublethal water deficit, the individual from which we took the leaf for WSD determination was exposed to considerable stress, and the leaf was damaged, even though we tried to take leaves healthy, vital and undamaged. The remaining WSD values for Populus alba were lower but indicate an adverse effect of the habitat on this species.
The WSD values for Diplotaxis muralis species are lower than for Populus alba but higher than for Calamagrostis epigejos. This fact is related to the fact that the Calamagrostis epigejos species is best adapted to the lack of water in the habitat, has scleromorphic leaves and can form an effective root system [32].
Tissue hydration is a necessary condition for the physiological activity of the leaves. The current water deficit depends on the balance between water intake and expenditure by the plant, so it is conditioned by many environmental and biological factors [25].
Samples of selected species were taken in July under relatively unfavourable humidity conditions in the range of 18.1–22.9%. However, at the last field sampling in September, the relative air humidity ranged from 41.1 to 60.8%.
4.2. Plant Utilisation in Landfill Remediation Strategies
From the obtained data we could state that all plant species were able to be inhabited and grow in the landfill waste, while each species had a different life strategy and way to adapt to this habitat with extreme conditions. Calamagrostis epigejos proved to be a plastic species, and in the selected habitat it was probably the best of all species for water shortages. Diplotaxis muralis showed the best adaptation, mainly in terms of the specific leaf area, the dry matter content in the leaves but also values of the water saturation deficit. Populus alba, although in the juvenile stage and affected by the lack of substrate moisture, was also able to adapt, colonize and create populations in habitats with adverse conditions. All three species belong to the same ecological group in terms of salinity [41]. They are tolerant to the concentration of readily soluble salts, especially sulphates, chlorides and carbonates of sodium, potassium, calcium and magnesium.
Despite the unfavourable conditions persisting in the landfill waste, all three species were able to create viable populations. Many plant species can accumulate heavy metals in their bodies and are therefore often sought after as indicators of contaminated areas or are used directly to revitalize and phytoextract heavy metals from soils [15]. Within the plant kingdom, about 500 species of higher plants are currently known to have hyperaccumulating properties [42]. Most of them can accumulate a specific element and can be used in phytoremediation methods. These species occur in 34 families, most of which belong to Brassicaceae family [42,43]. Diplotaxis muralis belongs to the family Brassicaceae, and even though it is not referred to as a hyperaccumulator, this species proved its ability to adapt to the high content of heavy metals in the substrate. In addition, the plasticity of species Populus alba and Calamagrostis epigejos allows them to grow in contaminated sites. These species are able to tolerate high levels of contaminants in the soil and partially accumulate them in their tissues [33,44]. Poplar is a sought-after species for the remediation of polluted sites due to its rapid growth, deep root system, large biomass production [45,46]. This species is suitable for soil rhizodegradation and phytoextraction of Cd, Ni, Fe, Zn and Pb [33,44,47,48].
Randelovic et al. [49] have been working directly with Calamagrostis epigejos. They concluded that its properties such as adaptation mechanism, phenotypic plasticity, genotypic variability, low nutrient requirements support its spread in this type of site and predestines it to indicate soil pollution and possible phytoremediation of contaminated areas. Its ability to colonize landfills after mining activities [50] plays an essential role in not spreading contaminants into the environment and crops. The species is suitable for remediation of soils polluted with heavy metals [48].
Phytocapping, as one of the phytoremediation technologies, is a new ecofriendly and cost-effective method working with overlapping landfill wastes with green naturally occurring plants. The main advantage of phytocapping is the reduction or complete stopping of heavy metal leaching into soil and groundwater. Plant biomass also slows down the spread of contaminants through the air and helps prevent erosion. Plants growing on landfill wastes are adapted to these conditions, and the concentration of heavy elements gradually decreases. Thanks to surface evaporation and plant transpiration, contaminated water is prevented from flowing down into the subsoil. Soil absorbs water and plants behave as “pumps” that can remove stored water [51,52,53]. The proposed phytocapping procedures recommend the use of hyperaccumulators and especially as the first step it is recommended to create a layer on contaminated soil, which will create a suitable environment for the plant roots [13]. For phytocapping, however, we can also use plants that themselves have the ability to adapt and grow on contaminated soils. If the landfill wastes are left without remediation methods for economic reasons, and natural vegetation is gradually created on it, we can talk about natural phytocapping.
5. Conclusions
The landfill waste (SW Slovakia) and its surroundings represent, from the environmental point of view, a territory that is severely disturbed, ecologically unresolved and left to the self-cleaning ability of the natural landscape. The waste containing heavy metals has contaminated a wide area, and the landfill waste greening is considered the most suitable method of reclamation. For this reason, the site was selected for field research and more detailed study, which focused on its natural vegetation.
The functional traits of three plant species that occur naturally at the site were evaluated, and the degree of adaptation of the plants to soil contaminated with heavy metals was determined. We focused on the species Populus alba, Diplotaxis muralis and Calamagrostis epigejos, which formed more numerous populations in this habitat. We evaluated leaf characteristics on selected individuals (specific leaf area, dry matter content in leaves, water saturation deficit and leaf water content). The highest values of the specific leaf area were determined for Diplotaxis muralis and the lowest in the case of Populus alba. On the contrary, Diplotaxis muralis showed the lowest values of leaf dry matter content, while the other two species, Populus alba and Calamagrostis epigejos, showed considerably higher values. The highest values of water saturation deficit were observed for Populus alba, Diplotaxis muralis had slightly lower values, and Calamagrostis epigejos showed the lowest values. Results of leaf water content revealed the best water management ability in the case of Diplotaxis muralis. This was confirmed by the significant correlation between specific leaf area and leaf water content of Diplotaxis muralis. We assume that this is related to the excellent plasticity of the species and its ability to adapt to adverse conditions.
Based on the investigated functional traits, which are closely related to the ability of plants to adapt to the contaminated environment, we can conclude that if it is not possible to dispose of the landfill wastes conventionally, then natural greening is a suitable solution. The natural plant cover can be formed by plants that are able to plastically respond to the high contents of heavy metals in the soil and can create strong and viable populations. These plant species reduce water percolation due to interception and evapotranspiration, prevent water and wind erosion of the soil and therefore reduce the spread of contamination to the environment.
Author Contributions
Conceptualization, I.V., M.Č. and T.O., methodology, I.V. and E.M.; formal analysis, T.O. and M.Č.; investigation, I.V., M.Č.; resources, E.M. and T.O.; data curation, M.Č.; writing—original draft preparation, M.Č.; writing—review and editing, M.Č.; visualization, T.O.; supervision, I.V. and E.M.; project administration, M.Č., I.V. and T.O.; funding acquisition, I.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Slovak Scientific Grant Agency VEGA projects 1/0712/20 and 2/0096/19.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gaggero, E.; Malandrino, M.; Fabbri, D.; Bordiglia, G.; Fusconi, A.; Mucciarelli, M.; Inaudi, P.; Calza, P. Uptake of Potentially Toxic Elements by Four Plant Species Suitable for Phytoremediation of Turin Urban Soils. Appl. Sci. 2020, 10, 3948. [Google Scholar] [CrossRef]
- Hu, Y.; Yu, Z.; Fang, X.; Zhang, W.; Liu, J.; Zhao, F. Influence of Mining and Vegetation Restoration on Soil Properties in the Eastern Margin of the Qinghai-Tibet Plateau. Int. J. Environ. Res. Public Health 2020, 17, 4288. [Google Scholar] [CrossRef] [PubMed]
- Pająk, M.; Błońska, E.; Szostak, M.; Gąsiorek, M.; Pietrzykowski, M.; Urban, O.; Derbis, P. Restoration of Vegetation in Relation to Soil Properties of Spoil Heap Heavily Contaminated with Heavy Metals. Water Air Soil Pollut. 2018, 229, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaeli, E.; Boltižiar, M.; Fazekašová, D.; Solár, V. The present character of the habitats on the industrial waste of Lúženec in the Sered’ (Slovak Republic). J. Int. Sci. Publ. Ecol. Saf. 2013, 7, 1313–2563. [Google Scholar]
- Edraki, M.; Baumgartl, T.; Manlapig, E.; Bradshaw, D.; Franks, D.M.; Moran, C.J. Designing mine tailings for better environmental, social and economic outcomes: A review of alternative approaches. J. Clean. Prod. 2014, 84, 411–420. [Google Scholar] [CrossRef]
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. ISRN Ecol. 2011, 2011, 402647. [Google Scholar] [CrossRef] [Green Version]
- Sharma, H.D.; Reddy, K.R. Geoenvironmental Engineering: Site Remediation, Waste Containment, and Emerging Waste Management Technologies; John Wiley & Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- García-Valero, A.; Martínez-Martínez, S.; Faz, A.; Rivera, J.; Acosta, J. Environmentally sustainable acid mine drainage remediation: Use of natural alkaline material. J. Water Process. Eng. 2020, 33, 101064. [Google Scholar] [CrossRef]
- Peppas, A.; Komnitsas, K.; Halikia, I. Use of organic covers for acid mine drainage control. Min. Eng. 2000, 13, 563–574. [Google Scholar] [CrossRef]
- Dermont, G.; Bergeron, M.; Mercier, G.; Richer-LaFLèche, M. Soil washing for metal removal: A review of physical/chemical technologies and field applications. J. Hazard. Mater. 2008, 152, 1–31. [Google Scholar] [CrossRef]
- Cameselle, C.; Pena, A. Enhanced electromigration and electro-osmosis for the remediation of an agricultural soil contaminated with multiple heavy metals. Process Saf. Environ. Prot. 2016, 104, 209–217. [Google Scholar] [CrossRef]
- Valenzuela, E.I.; García-Figueroa, A.C.; Amábilis-Sosa, L.E.; Molina-Freaner, F.E.; Pat-Espadas, A.M. Stabilization of potentially toxic elements contained in mine waste: A microbiological approach for the environmental management of mine tailings. J. Environ. Manag. 2020, 270, 110873. [Google Scholar] [CrossRef] [PubMed]
- Karaca, O.; Cameselle, C.; Reddy, K.R. Mine tailing disposal sites: Contamination problems, remedial options and phytocaps for sustainable remediation. Rev. Environ. Sci. Bio/Technol. 2018, 17, 205–228. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H. Phytoremediation. Annu. Rev. Plant Biol. 2005, 56, 15–39. [Google Scholar] [CrossRef] [PubMed]
- Salt, D.E.; Smith, R.D.; Raskin, I. Phytoremediation. Ann. Rev. Plant Biol. 1998, 49, 643–668. [Google Scholar] [CrossRef]
- Michaeli, E.; Boltižiar, M.; Solár, V.; Vilček, J.; Ivanová, M. Landfill Waste from the Production of Nickel and Cobalt in the Slovak Republic. Int. J. Met. Mater. Eng. 2016, 2, 126. [Google Scholar] [CrossRef]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Caudullo, G.; de Rigo, D. Populus alba in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publ. Off. EU: Luxembourg, 2016; pp. 134–135. [Google Scholar]
- Wild, J.; Kaplan, Z.; Danihelka, J.; Petřík, P.; Chytrý, M.; Novotný, P.; Rohn, M.; Šulc, V.; Brůna, J.; Chobot, K.; et al. Plant distribution data for the Czech Republic integrated in the Pladias database. Preslia 2019, 91, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Plants of the World Online. Available online: http://www.plantsoftheworldonline.org/taxon/urn:lsid:ipni.org:names:394433-1#distribution-map (accessed on 18 November 2020).
- Pladias—Databáze České Flóry a Vegetace. Available online: https://pladias.cz/taxon/data/Diplotaxis%20muralis (accessed on 15 November 2020).
- Clayton, W.D.; Govaerts, R.; Harman, K.T.; Williamson, H.; Vorontsova, M. World Checklist of Poaceae. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://wcsp.science.kew.org/ (accessed on 18 November 2020).
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Masarovičová, E.; Májeková, M.; Vykouková, I. Functional Traits and Plasticity of Plants. In Handbook of Photosynthesis, 3rd ed.; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 545–563. [Google Scholar]
- Perezharguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Garnier, E.; Shipley, B.; Roumet, C.; Laurent, G. A standardized protocol for the determination of specific leaf area and leaf dry matter content. Funct. Ecol. 2001, 15, 688–695. [Google Scholar] [CrossRef]
- Garnier, E.; Laurent, G. Leaf anatomy, specific mass and water content in congeneric annual and perennial grass species. New Phytol. 1994, 128, 725–736. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, T.; Skidmore, A.K.; Darvishzadeh, R.; Niemann, K.O.; Liu, J. Canopy leaf water content estimated using terrestrial LiDAR. Agric. For. Meteorol. 2017, 232, 152–162. [Google Scholar] [CrossRef]
- Pasqualotto, N.; Delegido, J.; Van Wittenberghe, S.; Verrelst, J.; Rivera, J.P.; Moreno, J. Retrieval of canopy water content of different crop types with two new hyperspectral indices: Water Absorption Area Index and Depth Water Index. Int. J. Appl. Earth Obs. Geoinf. 2018, 67, 69–78. [Google Scholar] [CrossRef]
- Mitrović, M.; Pavlović, P.; Lakušić, D.; Djurdjević, L.; Stevanović, B.; Kostić, O.; Gajić, G. The potential of Festuca rubra and Calamagrostis epigejos for the revegetation of fly ash deposits. Sci. Total Environ. 2008, 407, 338–347. [Google Scholar] [CrossRef]
- Rafati, M.; Khorasani, N.; Moattar, F.; Shirvany, A.; Moraghebi, F.; Hosseinzadeh, S. Phytoremediation potential of Populus alba and Morus alba for cadmium, chromuim and nickel absorption from polluted soil. Int. J. Environ. Res. 2011, 5, 961–970. [Google Scholar]
- Witkowski, E.T.F.; Lamont, B.B. Leaf specific mass confounds leaf density and thickness. Oecologia 1991, 88, 486–493. [Google Scholar] [CrossRef]
- Cunningham, S.A.; Summerhayes, B.; Westoby, M. Evolutionary divergences in leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecol. Monogr. 1999, 69, 569–588. [Google Scholar] [CrossRef]
- McIntyre, S.; Díaz, S.; Lavorel, S.; Cramer, W. Plant functional types and disturbance dynamics—Introduction. J. Veg. Sci. 1999, 10, 603–608. [Google Scholar] [CrossRef]
- van der Werf, A.; Geerts, R.H.E.M.; Jacobs, F.H.H.; Korevaar, H.; Oomes, M.J.M.; de Visser, W. The importance of relative growth rate and associated traits for competition between species during vegetation succession. In Inherent Variation in Plant Growth. Physiological Mechanisms and Ecological Consequences; Lambers, H., Poorter, H., van Vuuren, M.M.I., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 498–502. [Google Scholar]
- Grime, J.P.; Cornelissen, J.H.C.; Thompson, K.; Hodgson, J.G. Evidence of a Causal Connection between Anti-Herbivore Defence and the Decomposition Rate of Leaves. Oikos 1996, 77, 489–494. [Google Scholar] [CrossRef]
- Cingolani, A.M.; Cabido, M.; Gurvich, D.E.; Renison, D.; Díaz, S. Filtering processes in the assembly of plant communities: Are species presence and abundance driven by the same traits? J. Veg. Sci. 2007, 18, 911–920. [Google Scholar] [CrossRef]
- Slavík, B. Water Stress in Plants: Proceedings of a Symposium Held in Prague, September 30–October 4, 1963, 1st ed.; Springer: Amsterdam, The Netherlands, 1965. [Google Scholar]
- Chytrý, M.; Tichý, L.; Dřevojan, P.; Sádlo, J.; Zelený, D. Ellenberg-type indicator values for the Czech flora. Preslia 2018, 90, 83–103. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.; Nongkynrih, J.M. Metal hyperaccumulation and bioremediation. Biol. Plant. 2007, 51, 618–634. [Google Scholar] [CrossRef]
- Chaney, R.L.; Angle, J.S.; Broadhurst, C.L.; Peters, C.A.; Tappero, R.V.; Sparks, D.L. Improved understanding of hyperaccumulation yields commercial phytoextraction and phytomining technologies. J. Environ. Qual. 2007, 36, 1429–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghi, M.; Tognetti, R.; Monteforti, G.; Sebastiani, L. Responses of two poplar species (Populus alba and Populus x canadensis) to high copper concentrations. Environ. Exp. Bot. 2008, 62, 290–299. [Google Scholar] [CrossRef]
- Fischerová, Z.; Tlustoš, P.; Száková, J.; Šichorová, K. A comparison of phytoremediation capability of selected plant species for given trace elements. Environ. Pollut. 2006, 144, 93–100. [Google Scholar] [CrossRef]
- Castiglione, S.; Franchin, C.; Fossati, T.; Lingua, G.; Torrigiani, P.; Biondi, S. High zinc concentrations reduce rooting capacity and alter metallothionein gene expression in white poplar (Populus alba L. cv. Villafranca). Chemosphere 2007, 67, 1117–1126. [Google Scholar] [CrossRef]
- Madejon, P.; Maranon, T.; Murillo, J.M.; Robinson, B. White poplar (Populus alba) as a biomonitor of trace elements in contaminated riparian forests. Environ. Pollut. 2004, 132, 145–155. [Google Scholar] [CrossRef]
- Gorelova, S.V.; Frontasyeva, M.V. The use of higher plants in biomonitoring and environmental bioremediation. In Phytoremediation: Management of Environmental Contaminants; Ansari, A.A., Gill, S.S., Lanza, G.R., Newman, L., Eds.; Springer: Cham, Switzerland, 2017; pp. 103–155. [Google Scholar]
- Ranđelović, D.; JakovljeviĆ, K.; Jovanović, S. The application of Calamagrostis epigejos (L.) Roth. in phytoremediation technologies. In Phytoremediation Potential of Perennial Grasses; Pandey, V.C., Singh, D.P., Eds.; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 259–282. [Google Scholar]
- Mudrák, O.; Frouz, J.; Velichová, V. Understory vegetation in reclaimed and unreclaimed post-mining forest stands. Ecol. Eng. 2010, 36, 783–790. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, K.; Shah, M.P.; Kumar, M. Phytocapping: An eco-sustainable green technology for environmental pollution control. In Bioremediation for Environmental Sustainability; Kumar, V., Saxena, G., Shah, M.P., Eds.; Elsevier BV: Amsterdam, The Netherlands, 2021; pp. 481–491. [Google Scholar]
- Salt, M.; Yuen, S.T.S.; Ashwath, N.; Sun, J.; Benaud, P.; Zhu, G.X.; Jaksa, M.B.; Ghadiri, H.; Greenway, M.; Fourie, A.B. Phytocapping of landfills. In Solid Waste Landfilling: Concepts, Processes, Technology; Cossu, R., Stegmann, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 677–688. [Google Scholar]
- Khapre, A.; Kumar, S.; Rajasekaran, C. Phytocapping: An alternate cover option for municipal solid waste landfills. Environ. Technol. 2017, 40, 2242–2249. [Google Scholar] [CrossRef]
Figure 1.
Map of Slovakia with the landfill waste location.

Figure 2.
Actual view of the landfill.

Figure 3.
Populus alba growing in the landfill.

Figure 4.
Calamagrostis epigejos growing in the landfill.

Figure 5.
Diplotaxis muralis growing in the landfill.

Figure 6.
Specific leaf area (SLA) of investigated plant species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos).
Figure 6.
Specific leaf area (SLA) of investigated plant species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos).

Figure 7.
Dry matter content in the leaves (LDMC) of investigated species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos).
Figure 7.
Dry matter content in the leaves (LDMC) of investigated species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos).

Figure 8.
Water saturation deficit (WSD) in the leaves of investigated species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos).
Figure 8.
Water saturation deficit (WSD) in the leaves of investigated species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos).

Figure 9.
Leaf water content (LWC) of investigated species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos).
Figure 9.
Leaf water content (LWC) of investigated species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos).

Figure 10.
Correlations between a specific leaf area (SLA) and leaf water content (LWC) of the selected species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos) (values for SLA are given in cm2·g−1 and for LWC in g·g−1).
Figure 10.
Correlations between a specific leaf area (SLA) and leaf water content (LWC) of the selected species (PA—Populus alba, DM—Diplotaxis muralis, CE—Calamagrostis epigejos) (values for SLA are given in cm2·g−1 and for LWC in g·g−1).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Čierniková, M.; Vykouková, I.; Orfánus, T.; Masarovičová, E. Functional Traits of Plant Species Suitable for Revegetation of Landfill Waste from Nickel Smelter. Appl. Sci. 2021, 11, 658. https://doi.org/10.3390/app11020658
AMA Style
Čierniková M, Vykouková I, Orfánus T, Masarovičová E. Functional Traits of Plant Species Suitable for Revegetation of Landfill Waste from Nickel Smelter. Applied Sciences. 2021; 11(2):658. https://doi.org/10.3390/app11020658
Chicago/Turabian StyleČierniková, Malvína, Ivana Vykouková, Tomáš Orfánus, and Elena Masarovičová. 2021. "Functional Traits of Plant Species Suitable for Revegetation of Landfill Waste from Nickel Smelter" Applied Sciences 11, no. 2: 658. https://doi.org/10.3390/app11020658
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.